| 6-phosphofructo-2-kinase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 6-phosphofructo-2-kinase dimer, Human heart | |||||||||
| Identifiers | |||||||||
| EC no. | 2.7.1.105 | ||||||||
| CAS no. | 78689-77-7 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| Gene Ontology | AmiGO / QuickGO | ||||||||
| |||||||||
| 6PF2K | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 crystal structure of human liver 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase | |||||||||
| Identifiers | |||||||||
| Symbol | 6PF2K | ||||||||
| Pfam | PF01591 | ||||||||
| Pfam clan | CL0023 | ||||||||
| InterPro | IPR013079 | ||||||||
| PROSITE | PDOC00158 | ||||||||
| SCOP2 | 1bif / SCOPe / SUPFAM | ||||||||
| |||||||||
| 6-phosphofructo-2-kinase/fructose-bisphosphatase-2 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
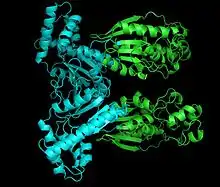 Structure of PFK2. Shown: kinase domain (cyan) and the phosphatase domain (green). | |||||||||||
| Identifiers | |||||||||||
| Symbol | 6PF2K | ||||||||||
| Pfam | PF01591 | ||||||||||
| InterPro | IPR013079 | ||||||||||
| PROSITE | PDOC00158 | ||||||||||
| SCOP2 | 1bif / SCOPe / SUPFAM | ||||||||||
| |||||||||||
| fructose-bisphosphatase-2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| Symbol | FBPase-2 | ||||||||
| Pfam | PF00316 | ||||||||
| InterPro | IPR028343 | ||||||||
| PROSITE | PDOC00114 | ||||||||
| |||||||||
Phosphofructokinase-2 (6-phosphofructo-2-kinase, PFK-2) or fructose bisphosphatase-2 (FBPase-2), is an enzyme indirectly responsible for regulating the rates of glycolysis and gluconeogenesis in cells. It catalyzes formation and degradation of a significant allosteric regulator, fructose-2,6-bisphosphate (Fru-2,6-P2) from substrate fructose-6-phosphate. Fru-2,6-P2 contributes to the rate-determining step of glycolysis as it activates enzyme phosphofructokinase 1 in the glycolysis pathway, and inhibits fructose-1,6-bisphosphatase 1 in gluconeogenesis.[1] Since Fru-2,6-P2 differentially regulates glycolysis and gluconeogenesis, it can act as a key signal to switch between the opposing pathways.[1] Because PFK-2 produces Fru-2,6-P2 in response to hormonal signaling, metabolism can be more sensitively and efficiently controlled to align with the organism's glycolytic needs.[2] This enzyme participates in fructose and mannose metabolism. The enzyme is important in the regulation of hepatic carbohydrate metabolism and is found in greatest quantities in the liver, kidney and heart. In mammals, several genes often encode different isoforms, each of which differs in its tissue distribution and enzymatic activity.[3] The family described here bears a resemblance to the ATP-driven phospho-fructokinases, however, they share little sequence similarity, although a few residues seem key to their interaction with fructose 6-phosphate.[4]
PFK-2 is known as the "bifunctional enzyme" because of its notable structure: though both are located on one protein homodimer, its two domains act as independently functioning enzymes.[5] One terminus serves as a kinase domain (for PFK-2) while the other terminus acts as a phosphatase domain (FBPase-2).[6]
In mammals, genetic mechanisms encode different PFK-2 isoforms to accommodate tissue specific needs. While general function remains the same, isoforms feature slight differences in enzymatic properties and are controlled by different methods of regulation; these differences are discussed below.[7]
Structure
The monomers of the bifunctional protein are clearly divided into two functional domains. The kinase domain is located on the N-terminal.[8] It consists of a central six-stranded β sheet, with five parallel strands and an antiparallel edge strand, surrounded by seven α helices.[6] The domain contains nucleotide-binding fold (nbf) at the C-terminal end of the first β-strand.[9] The PFK-2 domain appears to be closely related to the superfamily of mononucleotide binding proteins including adenylate cyclase.[10]
On the other hand, the phosphatase domain is located on the C-terminal.[11] It resembles the family of proteins that include phosphoglycerate mutases and acid phosphatases.[10][12] The domain has a mixed α/ β structure, with a six-stranded central β sheet, plus an additional α-helical subdomain that covers the presumed active site of the molecule.[6] Finally, the N-terminal region modulates PFK-2 and FBPase2 activities, and stabilizes the dimer form of the enzyme.[12][13]
While this central catalytic core remains conserved in all forms of PFK-2, slight structural variations exist in isoforms as a result of different amino acid sequences or alternative splicing.[14] With some minor exceptions, the size of PFK-2 enzymes is typically around 55 kDa.[1]
Researchers hypothesize that the unique bifunctional structure of this enzyme arose from a gene fusion event between a primordial bacterial PFK-1 and a primordial mutase/phosphatase.[15]
Function
This enzyme's main function is to synthesize or degrade allosteric regulator Fru-2,6-P2 in response to glycolytic needs of the cell or organism, as depicted in the accompanying diagram.

In enzymology, a 6-phosphofructo-2-kinase (EC 2.7.1.105) is an enzyme that catalyzes the chemical reaction:
- ATP + beta-D-fructose 6-phosphate ADP + beta-D-fructose 2,6-bisphosphate[16]

Thus, the kinase domain hydrolyzes ATP to phosphorylate the carbon-2 of fructose-6-phosphate, producing Fru-2,6-P2 and ADP. A phosphohistidine intermediate is formed within the reaction.[17]
- At the other terminal, the fructose-2,6-bisphosphate 2-phosphatase (EC 3.1.3.46) domain dephosphorylates Fru-2,6-P2 with the addition of water. This opposing chemical reaction is:
- beta-D-fructose 2,6-bisphosphate + H2O D-fructose 6-phosphate + phosphate[18]
Because of the enzyme's dual functions, it can be categorized into multiple families. Through categorization by the kinase reaction, this enzyme belongs to the family of transferases, specifically those transferring phosphorus-containing groups (phosphotransferases) with an alcohol group as acceptor.[16] On the other hand, the phosphatase reaction is characteristic of the family of hydrolases, specifically those acting on phosphoric monoester bonds.[18]
Regulation
In almost all isoforms, PFK-2 undergoes covalent modification through phosphorylation/dephosphorylation based on the cell's hormonal signaling. Phosphorylation of a specific residue may prompt a shift that stabilizes either kinase or phosphatase domain function. This regulation signal thus controls whether F-2,6-P2 will be synthesized or degraded.[19]
Furthermore, the allosteric regulation of PFK2 is very similar to the regulation of PFK1.[20] High levels of AMP or phosphate group signifies a low energy charge state and thus stimulates PFK2. On the other hand, a high concentration of phosphoenolpyruvate (PEP) and citrate signifies that there is a high level of biosynthetic precursor and hence inhibits PFK2. Unlike PFK1, PFK2 is not affected by ATP concentration.[21]
Isozymes
Protein isozymes are enzymes that catalyze the same reaction but are encoded with different amino acid sequences and as such, display slight differences in protein characteristics. In humans, the four genes that encode phosphofructokinase 2 proteins include PFKFB-1, PFKFB2, PFKFB3 and PFKFB4.[5]
Multiple mammalian isoforms of the protein have been reported to date, difference rising by either the transcription of different enzymes or alternative splicing.[22][23][24] While the structural core that catalyzes the PFK-2/FBPase-2 reaction is highly conserved across isoforms, the major differences arise from highly variable flanking sequences in the isoform amino and carboxyl terminals.[14] Because these areas often contain phosphorylation sites, changes in amino acid composition or terminal length may result in vastly different enzyme kinetics and characteristics.[1][14] Each variant differs in their primary tissue of expression, response to protein kinase regulation, and ratio of kinase/phosphatase domain activity.[25] While multiple types of isozymes may consist in a tissue, isozymes are identified by their primary tissue expression and tissue of discovery below.[26]
PFKB1: Liver, muscle, and fetal
| 6-phosphofructo-2-kinase: PFKB1 | |||||||||
|---|---|---|---|---|---|---|---|---|---|
Crystal structure of human liver 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase | |||||||||
| Identifiers | |||||||||
| EC no. | 2.7.1.105 | ||||||||
| CAS no. | 78689-77-7 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| Gene Ontology | AmiGO / QuickGO | ||||||||
| |||||||||
Located on the X chromosome, this gene is the most well-known of the four genes particularly because it encodes the highly researched liver enzyme.[22] Variable mRNA splicing of PFKB1 yields three different promoters (L, M and F) and therefore, three tissue-specific variants that differ in regulation:[27]
- L-Type: liver tissue
- Insulin activates liver PFK-2 function to indicate a high abundance of blood glucose is available for glycolysis. Insulin activates a protein phosphatase which dephosphorylates the PFK-2 complex and causes favored PFK-2 activity. PFK-2 then increases production of F-2,6-P2. As this product allosterically activates PFK-1, it activates glycolysis and inhibits gluconeogenesis.[28]
- In contrast, glucagon increases FBPase-2 activity. At low blood glucose concentrations, glucagon triggers a cAMP signal cascade and in turn, Protein Kinase A (PKA) phosphorylates Serine 32 near the N-terminus. This inactivates the bifunctional enzyme's ability to act as a kinase and stabilizes the phosphatase activity. Therefore, glucagon decreases concentrations of F-2,6-P2, slows rates of glycolysis, and stimulates the gluconeogenesis pathway.[29][30]

- M-Type: skeletal muscle tissue; F-Type: fibroblast and fetal tissue[31]
- In contrast to most other PFK-2 tissues, PFK-2 in both skeletal muscle and fetal tissue is solely regulated by concentrations of Fructose-6-phosphate. Within their first exon, there are no regulatory sites that require phosphorylation/dephosphorylation to provoke a change in function. High concentrations of F-6-P will activate kinase function and increase rates of glycolysis, whereas low concentrations of F-6-P will stabilize phosphatase action.[27]
| 6-phosphofructo-2-kinase: PFKB2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|
6-phosphofructo-2-kinase dimer, Human heart tissue | |||||||||
| Identifiers | |||||||||
| EC no. | 2.7.1.105 | ||||||||
| CAS no. | 78689-77-7 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| Gene Ontology | AmiGO / QuickGO | ||||||||
| |||||||||
PFKB2: Cardiac (H-Type)
The PFKB2 gene is located on chromosome 1.[32] When greater concentrations of adrenaline and/or insulin hormone are circulated, a Protein Kinase A pathway is activated which phosphorylates either Serine 466 or Serine 483 in the C-terminus.[3] Alternatively, Protein Kinase B may also phosphorylate these regulatory sites, which are part of the FBPase-2 domain.[33] When this serine residue is phosphorylated, FBPase-2 function is inactivated and greater PFK-2 activity is stabilized.[27]
PFKB3: Brain, placental, and inducible
PFKB3 is located on chromosome 10 and transcribes two major isoforms, inducible type and ubiquitous type.[34] These forms differ in alternative splicing of Exon 15 in their C-terminus.[35] However, they are similar in that for both, glucagon activates a cyclic AMP pathway; this results in Protein Kinase A, Protein Kinase C, or AMP-activated Protein Kinase phosphorylating a regulatory residue on Serine 461 in the C-terminus to stabilize PFK-2 kinase function.[36] Furthermore, both isoforms transcribed from this gene are noted for having a particularly high, dominant rate of kinase activity as indicated by a kinase/phosphatase activity ratio of 700 (whereas the liver, heart, and testis isozymes respectively have PFK-2/FBPase-2 ratios of 1.5, 80, and 4).[37] Therefore, PFKB3 in particular consistently produces large amounts of F-2,6-P2 and sustains high rates of glycolysis.[37][38]
- I-Type: Inducible
- U-Type: Ubiquitous;[40] also known as placental[41] or brain[42][43]
- Though discovered separately in the placental, pancreatic-β-islet, or brain tissues, the various isoforms appear identical.[21] The tissues it was discovered in all require great energy to function, which may explain PFKB3's advantage of such high kinase-phosphatase activity ratio.[37][44]
- The brain isoform in particular has lengthy N- and C-terminus regions such that this type is almost twice as large as the typical PFK-2, at around 110 kDa.[45]
PFKB4: Testis (T-Type)
Gene PFKB4, located on chromosome 3, expresses PFK-2 in human testis tissue.[46] PFK-2 enzymes encoded by PFK-4 are comparable to the liver enzyme in size at around 54kDa, and like the muscle tissue, do not contain a protein kinase phosphorylation site.[40] While less research has clarified regulation mechanisms for this isoform, studies have confirmed that modification from multiple transcription factors in the 5' flanking region regulates the amount of PFK-2 expression in developing testis tissue.[26] This isoform has been particularly implicated as being modified and hyper-expressed for prostate cancer cell survival.[47]

Clinical significance
Because this enzyme family maintains rates of glycolysis and gluconeogenesis, it presents great potential for therapeutic action for control of metabolism particularly in diabetes and cancer cells.[6][25] Data also demonstrates that all of the PFK-2 genes (although the PFKB3 gene response remains the most drastic) were activated by limitations in oxygen.[48] The control of PFK-2/FBP-ase2 activity was found to be linked to heart functioning, particularly for ischemia, and the control against hypoxia.[49] Researchers hypothesize that this responsive characteristic of the PFK-2 genes may be a strong, evolutionary physiological adaptation.[48] However, many human cancer cell types (including leukemia, lung, breast, colon, pancreatic, and ovarian cancers) demonstrate over-expression of PFK3 and/or PFK4; this change in metabolism likely plays a role in the Warburg effect.[25][50]
Lastly, the Pfkfb2 gene encoding PFK2/FBPase2 protein is linked to the predisposition to schizophrenia.[51]
References
- 1 2 3 4 Kurland IJ, Pilkis SJ (June 1995). "Covalent control of 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase: insights into autoregulation of a bifunctional enzyme". Protein Science. 4 (6): 1023–37. doi:10.1002/pro.5560040601. PMC 2143155. PMID 7549867.
- ↑ Lenzen S (May 2014). "A fresh view of glycolysis and glucokinase regulation: history and current status". The Journal of Biological Chemistry. 289 (18): 12189–94. doi:10.1074/jbc.R114.557314. PMC 4007419. PMID 24637025.
- 1 2 Heine-Suñer D, Díaz-Guillén MA, Lange AJ, Rodríguez de Córdoba S (May 1998). "Sequence and structure of the human 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase heart isoform gene (PFKFB2)". European Journal of Biochemistry. 254 (1): 103–10. doi:10.1046/j.1432-1327.1998.2540103.x. PMID 9652401.
- ↑ Wang X, Deng Z, Kemp RG (September 1998). "An essential methionine residue involved in substrate binding by phosphofructokinases". Biochem. Biophys. Res. Commun. 250 (2): 466–8. doi:10.1006/bbrc.1998.9311. PMID 9753654.
- 1 2 Rider MH, Bertrand L, Vertommen D, Michels PA, Rousseau GG, Hue L (August 2004). "6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase: head-to-head with a bifunctional enzyme that controls glycolysis". The Biochemical Journal. 381 (Pt 3): 561–79. doi:10.1042/BJ20040752. PMC 1133864. PMID 15170386.
- 1 2 3 4 Hasemann CA, Istvan ES, Uyeda K, Deisenhofer J (September 1996). "The crystal structure of the bifunctional enzyme 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase reveals distinct domain homologies". Structure. 4 (9): 1017–29. doi:10.1016/S0969-2126(96)00109-8. PMID 8805587.
- ↑ Atsumi T, Nishio T, Niwa H, Takeuchi J, Bando H, Shimizu C, Yoshioka N, Bucala R, Koike T (December 2005). "Expression of inducible 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase/PFKFB3 isoforms in adipocytes and their potential role in glycolytic regulation". Diabetes. 54 (12): 3349–57. doi:10.2337/diabetes.54.12.3349. PMID 16306349.
- ↑ Kurland I, Chapman B, Lee YH, Pilkis S (August 1995). "Evolutionary reengineering of the phosphofructokinase active site: ARG-104 does not stabilize the transition state in 6-phosphofructo-2-kinase". Biochemical and Biophysical Research Communications. 213 (2): 663–72. doi:10.1006/bbrc.1995.2183. PMID 7646523.
- ↑ Walker JE, Saraste M, Runswick MJ, Gay NJ (1982). "Distantly related sequences in the alpha- and beta-subunits of ATP synthase, myosin, kinases and other ATP-requiring enzymes and a common nucleotide binding fold". The EMBO Journal. 1 (8): 945–51. doi:10.1002/j.1460-2075.1982.tb01276.x. PMC 553140. PMID 6329717.
- 1 2 Jedrzejas MJ (2000). "Structure, function, and evolution of phosphoglycerate mutases: comparison with fructose-2,6-bisphosphatase, acid phosphatase, and alkaline phosphatase". Progress in Biophysics and Molecular Biology. 73 (2–4): 263–87. doi:10.1016/S0079-6107(00)00007-9. PMID 10958932.
- ↑ Li L, Lin K, Pilkis J, Correia JJ, Pilkis SJ (October 1992). "Hepatic 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase. The role of surface loop basic residues in substrate binding to the fructose-2,6-bisphosphatase domain". The Journal of Biological Chemistry. 267 (30): 21588–94. doi:10.1016/S0021-9258(19)36651-7. PMID 1328239.
- 1 2 Stryer L, Berg JM, Tymoczko JL (2008). "The Balance Between Glycolysis and Gluconeogenesis in the Liver Is Sensitive to Blood-Glucose Concentration". Biochemistry (Looseleaf). San Francisco: W. H. Freeman. pp. 466–467. ISBN 978-1-4292-3502-0.
- ↑ Tominaga N, Minami Y, Sakakibara R, Uyeda K (July 1993). "Significance of the amino terminus of rat testis fructose-6-phosphate, 2-kinase:fructose-2,6-bisphosphatase". The Journal of Biological Chemistry. 268 (21): 15951–7. doi:10.1016/S0021-9258(18)82344-4. PMID 8393455.
- 1 2 3 El-Maghrabi MR, Noto F, Wu N, Manes N (September 2001). "6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase: suiting structure to need, in a family of tissue-specific enzymes". Current Opinion in Clinical Nutrition and Metabolic Care. 4 (5): 411–8. doi:10.1097/00075197-200109000-00012. PMID 11568503. S2CID 6638455.
- ↑ Bazan JF, Fletterick RJ, Pilkis SJ (December 1989). "Evolution of a bifunctional enzyme: 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase". Proceedings of the National Academy of Sciences of the United States of America. 86 (24): 9642–6. Bibcode:1989PNAS...86.9642B. doi:10.1073/pnas.86.24.9642. PMC 298557. PMID 2557623.
- 1 2 "ENZYME entry 2.7.1.105". enzyme.expasy.org. Retrieved 2018-03-24.
- ↑ "6-phosphofructo-2-kinase (IPR013079)". InterPro. EMBL-EBI. Retrieved 2018-03-25.
- 1 2 "ENZYME entry 3.1.3.46". enzyme.expasy.org. Retrieved 2018-03-25.
- ↑ Okar DA, Manzano A, Navarro-Sabatè A, Riera L, Bartrons R, Lange AJ (January 2001). "PFK-2/FBPase-2: maker and breaker of the essential biofactor fructose-2,6-bisphosphate". Trends in Biochemical Sciences. 26 (1): 30–5. doi:10.1016/S0968-0004(00)01699-6. PMID 11165514.
- ↑ Van Schaftingen E, Hers HG (August 1981). "Phosphofructokinase 2: the enzyme that forms fructose 2,6-bisphosphate from fructose 6-phosphate and ATP". Biochemical and Biophysical Research Communications. 101 (3): 1078–84. doi:10.1016/0006-291X(81)91859-3. PMID 6458291.
- 1 2 Ros S, Schulze A (February 2013). "Balancing glycolytic flux: the role of 6-phosphofructo-2-kinase/fructose 2,6-bisphosphatases in cancer metabolism". Cancer & Metabolism. 1 (1): 8. doi:10.1186/2049-3002-1-8. PMC 4178209. PMID 24280138.
- 1 2 Darville MI, Crepin KM, Hue L, Rousseau GG (September 1989). "5' flanking sequence and structure of a gene encoding rat 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase". Proceedings of the National Academy of Sciences of the United States of America. 86 (17): 6543–7. Bibcode:1989PNAS...86.6543D. doi:10.1073/pnas.86.17.6543. PMC 297880. PMID 2549541.
- ↑ Tsuchiya Y, Uyeda K (May 1994). "Bovine heart fructose 6-P,2-kinase:fructose 2,6-bisphosphatase mRNA and gene structure". Archives of Biochemistry and Biophysics. 310 (2): 467–74. doi:10.1006/abbi.1994.1194. PMID 8179334.
- ↑ Sakata J, Abe Y, Uyeda K (August 1991). "Molecular cloning of the DNA and expression and characterization of rat testes fructose-6-phosphate,2-kinase:fructose-2,6-bisphosphatase". The Journal of Biological Chemistry. 266 (24): 15764–70. doi:10.1016/S0021-9258(18)98475-9. PMID 1651918.
- 1 2 3 Novellasdemunt L, Tato I, Navarro-Sabate A, Ruiz-Meana M, Méndez-Lucas A, Perales JC, Garcia-Dorado D, Ventura F, Bartrons R, Rosa JL (April 2013). "Akt-dependent activation of the heart 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase (PFKFB2) isoenzyme by amino acids". The Journal of Biological Chemistry. 288 (15): 10640–51. doi:10.1074/jbc.M113.455998. PMC 3624444. PMID 23457334.
- 1 2 Gómez M, Manzano A, Navarro-Sabaté A, Duran J, Obach M, Perales JC, Bartrons R (January 2005). "Specific expression of pfkfb4 gene in spermatogonia germ cells and analysis of its 5'-flanking region". FEBS Letters. 579 (2): 357–62. doi:10.1016/j.febslet.2004.11.096. PMID 15642344. S2CID 33170865.
- 1 2 3 Salway JG (2017). Metabolism at a Glance. Wiley-Blackwell. ISBN 978-0-470-67471-0.
- ↑ Hue L, Rider MH, Rousseau GG (1990). "Fructose-2,6-bisphosphate in extra hepatic tissues". In Pilkis SJ (ed.). Fructose-2,6-bisphosphate. Boca Raton, Fla.: CRC Press. pp. 173–193. ISBN 978-0-8493-4795-5.
- ↑ Pilkis SJ, el-Maghrabi MR, Claus TH (1988). "Hormonal regulation of hepatic gluconeogenesis and glycolysis". Annual Review of Biochemistry. 57: 755–83. doi:10.1146/annurev.bi.57.070188.003543. PMID 3052289.
- ↑ Marker AJ, Colosia AD, Tauler A, Solomon DH, Cayre Y, Lange AJ, el-Maghrabi MR, Pilkis SJ (April 1989). "Glucocorticoid regulation of hepatic 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase gene expression". The Journal of Biological Chemistry. 264 (12): 7000–4. doi:10.1016/S0021-9258(18)83531-1. PMID 2540168.
- ↑ Cosin-Roger J, Vernia S, Alvarez MS, Cucarella C, Boscá L, Martin-Sanz P, Fernández-Alvarez AJ, Casado M (February 2013). "Identification of a novel Pfkfb1 mRNA variant in rat fetal liver". Biochemical and Biophysical Research Communications. 431 (1): 36–40. doi:10.1016/j.bbrc.2012.12.109. hdl:11336/19538. PMID 23291237.
- ↑ Darville MI, Chikri M, Lebeau E, Hue L, Rousseau GG (August 1991). "A rat gene encoding heart 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase". FEBS Letters. 288 (1–2): 91–4. doi:10.1016/0014-5793(91)81009-W. PMID 1652483. S2CID 34116121.
- ↑ Marsin AS, Bertrand L, Rider MH, Deprez J, Beauloye C, Vincent MF, Van den Berghe G, Carling D, Hue L (October 2000). "Phosphorylation and activation of heart PFK-2 by AMPK has a role in the stimulation of glycolysis during ischaemia". Current Biology. 10 (20): 1247–55. doi:10.1016/S0960-9822(00)00742-9. PMID 11069105. S2CID 7920767.
- ↑ Riera L, Manzano A, Navarro-Sabaté A, Perales JC, Bartrons R (April 2002). "Insulin induces PFKFB3 gene expression in HT29 human colon adenocarcinoma cells". Biochimica et Biophysica Acta (BBA) - Molecular Cell Research. 1589 (2): 89–92. doi:10.1016/S0167-4889(02)00169-6. PMID 12007784.
- ↑ Marsin AS, Bouzin C, Bertrand L, Hue L (August 2002). "The stimulation of glycolysis by hypoxia in activated monocytes is mediated by AMP-activated protein kinase and inducible 6-phosphofructo-2-kinase". The Journal of Biological Chemistry. 277 (34): 30778–83. doi:10.1074/jbc.M205213200. PMID 12065600.
- 1 2 3 Sakakibara R, Kato M, Okamura N, Nakagawa T, Komada Y, Tominaga N, Shimojo M, Fukasawa M (July 1997). "Characterization of a human placental fructose-6-phosphate, 2-kinase/fructose-2,6-bisphosphatase". Journal of Biochemistry. 122 (1): 122–8. doi:10.1093/oxfordjournals.jbchem.a021719. PMID 9276680.
- ↑ Manes NP, El-Maghrabi MR (June 2005). "The kinase activity of human brain 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase is regulated via inhibition by phosphoenolpyruvate". Archives of Biochemistry and Biophysics. 438 (2): 125–36. doi:10.1016/j.abb.2005.04.011. PMID 15896703.
- ↑ Chesney J, Mitchell R, Benigni F, Bacher M, Spiegel L, Al-Abed Y, Han JH, Metz C, Bucala R (March 1999). "An inducible gene product for 6-phosphofructo-2-kinase with an AU-rich instability element: role in tumor cell glycolysis and the Warburg effect". Proceedings of the National Academy of Sciences of the United States of America. 96 (6): 3047–52. Bibcode:1999PNAS...96.3047C. doi:10.1073/pnas.96.6.3047. PMC 15892. PMID 10077634.
- 1 2 Manzano A, Rosa JL, Ventura F, Pérez JX, Nadal M, Estivill X, Ambrosio S, Gil J, Bartrons R (1998). "Molecular cloning, expression, and chromosomal localization of a ubiquitously expressed human 6-phosphofructo-2-kinase/ fructose-2, 6-bisphosphatase gene (PFKFB3)". Cytogenetics and Cell Genetics. 83 (3–4): 214–7. doi:10.1159/000015181. PMID 10072580. S2CID 23221556.
- ↑ Sakai A, Kato M, Fukasawa M, Ishiguro M, Furuya E, Sakakibara R (March 1996). "Cloning of cDNA encoding for a novel isozyme of fructose 6-phosphate, 2-kinase/fructose 2,6-bisphosphatase from human placenta". Journal of Biochemistry. 119 (3): 506–11. doi:10.1093/oxfordjournals.jbchem.a021270. PMID 8830046.
- ↑ Ventura F, Ambrosio S, Bartrons R, el-Maghrabi MR, Lange AJ, Pilkis SJ (April 1995). "Cloning and expression of a catalytic core bovine brain 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase". Biochemical and Biophysical Research Communications. 209 (3): 1140–8. doi:10.1006/bbrc.1995.1616. PMID 7733968.
- ↑ Bando H, Atsumi T, Nishio T, Niwa H, Mishima S, Shimizu C, Yoshioka N, Bucala R, Koike T (August 2005). "Phosphorylation of the 6-phosphofructo-2-kinase/fructose 2,6-bisphosphatase/PFKFB3 family of glycolytic regulators in human cancer". Clinical Cancer Research. 11 (16): 5784–92. doi:10.1158/1078-0432.CCR-05-0149. PMID 16115917.
- ↑ Riera L, Obach M, Navarro-Sabaté A, Duran J, Perales JC, Viñals F, Rosa JL, Ventura F, Bartrons R (August 2003). "Regulation of ubiquitous 6-phosphofructo-2-kinase by the ubiquitin-proteasome proteolytic pathway during myogenic C2C12 cell differentiation". FEBS Letters. 550 (1–3): 23–9. doi:10.1016/S0014-5793(03)00808-1. PMID 12935880. S2CID 41726316.
- ↑ Ventura F, Rosa JL, Ambrosio S, Pilkis SJ, Bartrons R (September 1992). "Bovine brain 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase. Evidence for a neural-specific isozyme". The Journal of Biological Chemistry. 267 (25): 17939–43. doi:10.1016/S0021-9258(19)37133-9. hdl:2445/177133. PMID 1325453.
- ↑ Manzano A, Pérez JX, Nadal M, Estivill X, Lange A, Bartrons R (March 1999). "Cloning, expression and chromosomal localization of a human testis 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase gene". Gene. 229 (1–2): 83–9. doi:10.1016/S0378-1119(99)00037-2. PMID 10095107.
- ↑ Ros S, Santos CR, Moco S, Baenke F, Kelly G, Howell M, Zamboni N, Schulze A (April 2012). "Functional metabolic screen identifies 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 4 as an important regulator of prostate cancer cell survival". Cancer Discovery. 2 (4): 328–43. doi:10.1158/2159-8290.CD-11-0234. PMID 22576210.
- 1 2 Minchenko, O., Opentanova, I., & Caro, J. (2003). Hypoxic regulation of the 6‐phosphofructo‐2‐kinase/fructose‐2, 6‐bisphosphatase gene family (PFKFB‐1–4) expression in vivo. FEBS Letters, 554(3), 264-270.
- ↑ Wang Q, Donthi RV, Wang J, Lange AJ, Watson LJ, Jones SP, Epstein PN (June 2008). "Cardiac phosphatase-deficient 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase increases glycolysis, hypertrophy, and myocyte resistance to hypoxia". American Journal of Physiology. Heart and Circulatory Physiology. 294 (6): H2889–97. doi:10.1152/ajpheart.91501.2007. PMC 4239994. PMID 18456722.
- ↑ Minchenko OH, Opentanova IL, Ogura T, Minchenko DO, Komisarenko SV, Caro J, Esumi H (2005). "Expression and hypoxia-responsiveness of 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase 4 in mammary gland malignant cell lines". Acta Biochimica Polonica. 52 (4): 881–8. doi:10.18388/abp.2005_3402. PMID 16025159.
- ↑ Stone WS, Faraone SV, Su J, Tarbox SI, Van Eerdewegh P, Tsuang MT (May 2004). "Evidence for linkage between regulatory enzymes in glycolysis and schizophrenia in a multiplex sample". American Journal of Medical Genetics. Part B, Neuropsychiatric Genetics. 127B (1): 5–10. doi:10.1002/ajmg.b.20132. PMID 15108172. S2CID 2420843.
- Van Schaftingen E, Hers HG (1981). "Phosphofructokinase 2: the enzyme that forms fructose 2,6-bisphosphate from fructose 6-phosphate and ATP". Biochem. Biophys. Res. Commun. 101 (3): 1078–84. doi:10.1016/0006-291X(81)91859-3. PMID 6458291.
External links
- Fructose+2,6-bisphosphatase at the U.S. National Library of Medicine Medical Subject Headings (MeSH)
- 6-phosphofructokinase of Arabidopsis thaliana at genome.jp
This article incorporates text from the public domain Pfam and InterPro IPR013079