| Ubiquinol oxidase (electrogenic, proton-motive force generating; Cytochrome bd) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
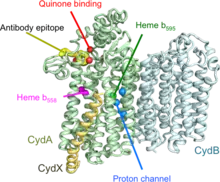 Predicted structure of E. coli Cytochrome bd-1 | |||||||||
| Identifiers | |||||||||
| EC no. | 7.1.1.7 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| |||||||||
Cytochrome d, previously known as cytochrome a2, is a name for all cytochromes (electron-transporting heme proteins) that contain heme D as a cofactor. Two unrelated classes of cytochrome d are known: Cytochrome bd, an enzyme that generates a charge across the membrane so that protons will move,[1] and cytochrome cd1 (NirS; SCOP b.70.2), a nitrite reductase.[2]
Cytochrome bd is found in plenty of aerobic bacteria, especially when it has grown with a limited oxygen supply. Compared to other terminal oxidases, it is notable for its high oxygen affinity and resistance to cyanide poisoning. It has a group of very similar relatives that do not use heme D, known as cyanide insensitive oxidases (CIOs).[3]
Function

Cytochrome d is, as other proteins of its family, a membrane-bound hemeprotein, but unlike cytochromes a and b, cytochrome D has a heme D instead of a heme A or heme B group.[4]
Cytochrome d is part of the cytochrome bd terminal oxidase which catalyse the two electron oxidation of ubiquinol. This process is an oxidative phosphorylation that oxidizes the ubiquinol-8 to ubiquinone. The chemical reaction followed by this process is:
- Ubiquinol-8 + O2 → Ubiquinone-8 + H2O[5]
By a similar reaction, it also catalyses the reduction of oxygen to water, which involves 4 electrons.
As a terminal oxidase, the reaction generates a proton motive force:
- 2 ubiquinol[inner membrane] + O2 + 4 H+[cytoplasm] → 2 ubiquinone[inner membrane] + 2 H2O + 4 H+[periplasm]
Some members of the family may accept or prefer other electron-transporting quinols such as menaquinol or plastoquinol in lieu of ubiquinol.[3]
Structure
Cytochrome bd (OPM family 805) is a tri-heme oxidase as it is compound by cytochromes b558, b595 and d. Its main function is the reduction of O2 to H2O. It is thought that it uses a di-heme active site, which is formed by the hemes of cytochromes b595 and d. These two cytochromes are considered high-spin complexes, what is directly related to the electrons' spin. While other respiratory terminal oxidases which catalyze that same reaction have a heme-copper active site and use a proton pump, cytochrome bd has an active site with iron instead of copper and need no proton pump as they can produce a proton-motion force themselves.[6] They are embedded in the bacterial cytoplasmic bilayer and serve as terminal oxidases in the respiratory chain.[7]
The oxidases tend to have two or three subunits. Subunits 1 (InterPro: IPR003317) and 2 (InterPro: IPR002585) are predicted to have pseudo-symmetry, and are sufficient to bind the two heme b molecules.[8] Some proteobacterial assemblies require a third subunit (InterPro: IPR012994) to bind heme d; others do not.[9]
The high-resolution structure heterotrimeric Cytochromes bd from Geobacillus species has been determined (PDB: 5IR6, 5DOQ). The third subunit does not share sequence homology with the third subunit proteobacteria, but does come into the assemblies at a similar position.[10]
Occurrence
Escherichia coli
E. coli possess two sets of Cytochrome bd.[7] The bd-I complex (CydABX) is a heterotrimer, while the bd-II complex (AppCB) is a heterodimer. There is an AppX gene that may correspond to a subunit 3 for AppCB.[9]
The ability of bd-II to generate a proton motive force is a matter of recent debate, putting it under the nonelectrogenic Ubiquinol oxidase (H+-transporting) in some categorizations.[11]
Azotobacter vinelandii
Azotobacter vinelandii is a nitrogen-fixing bacteria which is known by its high respiratory rate among aerobic organisms. Some physiological studies postulate that cytochrome d functions as a terminal oxidase in the membranes of this organism, taking part in the electron transport system. The studies characterized the different genes in the two subunits (Q09049, C1DEL1; third subunit C1DEL0). A very extensive homology with CydAB of the E. coli was found in these studies.[12]
Spectra
Generally, in protein complexes, cytochrome D gives an absorption band of approximately 636 nm or 638 nm, depending on the cytochrome d form. If it is oxidized, the band has a length of 636 nm, and a 638 nm length if it is reduced. It is commonly associated to certain prosthetic groups when found in multiple subunit complexes. Detecting cytochrome d as Fe(II) pyridine alkaline hemachrome is very difficult because the stability under these conditions is limited. If cytochrome d is pulled out of the protein complex (as heme D) and placed in ether containing from 1 to 5 % of HCl, it gives a different absorption band (603 nm, in the oxidized form).[2]
References
- ↑ EC 7.1.1.7
- 1 2 "Nomenclature Committee of the International Union of Biochemistry (NC-IUB). Nomenclature of electron-transfer proteins. Recommendations 1989" (PDF). European Journal of Biochemistry. 200 (3): 599–611. September 1991. doi:10.1111/j.1432-1033.1991.tb16223.x. PMID 1655423.
- 1 2 Borisov VB, Gennis RB, Hemp J, Verkhovsky MI (November 2011). "The cytochrome bd respiratory oxygen reductases". Biochimica et Biophysica Acta (BBA) - Bioenergetics. 1807 (11): 1398–413. doi:10.1016/j.bbabio.2011.06.016. PMC 3171616. PMID 21756872.
- ↑ Belevich I, Borisov VB, Konstantinov AA, Verkhovsky MI (August 2005). "Oxygenated complex of cytochrome bd from Escherichia coli: stability and photolability". FEBS Letters. 579 (21): 4567–70. doi:10.1016/j.febslet.2005.07.011. PMID 16087180. S2CID 36465802.
- ↑ Cytochrome d ubiquinol oxidase subunit 1
- ↑ Borisov VB, Verkhovsky MI (January 2013). "Accommodation of CO in the di-heme active site of cytochrome bd terminal oxidase from Escherichia coli". Journal of Inorganic Biochemistry. 118: 65–7. doi:10.1016/j.jinorgbio.2012.09.016. PMID 23123340.
- 1 2 Michael J. Miller, Robert B. Gennis. The Cytochrome d Complex Is a Coupling Site in the Aerobic Respiratory Chain of Escherichia coli. The Journal of Biological Chemistry Vol.260 No.26 (1985)
- ↑ Ovchinnikov S, Kinch L, Park H, Liao Y, Pei J, Kim DE, Kamisetty H, Grishin NV, Baker D (September 2015). "Large-scale determination of previously unsolved protein structures using evolutionary information". eLife. 4: e09248. doi:10.7554/eLife.09248. PMC 4602095. PMID 26335199.
- 1 2 Escherichia coli K-12 substr. MG1655 Transporter: cytochrome bd-I terminal oxidase
- ↑ Safarian S, Rajendran C, Müller H, Preu J, Langer JD, Ovchinnikov S, Hirose T, Kusumoto T, Sakamoto J, Michel H (April 2016). "Structure of a bd oxidase indicates similar mechanisms for membrane-integrated oxygen reductases". Science. 352 (6285): 583–6. Bibcode:2016Sci...352..583S. doi:10.1126/science.aaf2477. PMC 5515584. PMID 27126043.
- ↑ Borisov VB, Murali R, Verkhovskaya ML, Bloch DA, Han H, Gennis RB, Verkhovsky MI (October 2011). "Aerobic respiratory chain of Escherichia coli is not allowed to work in fully uncoupled mode". Proceedings of the National Academy of Sciences of the United States of America. 108 (42): 17320–4. Bibcode:2011PNAS..10817320B. doi:10.1073/pnas.1108217108. PMC 3198357. PMID 21987791.
- ↑ Jones CW, Redfearn ER. The cytochrome system of Azotobacter vinelandii. Biochim Biophys Acta. 1967 Sep 6;143(2):340–353