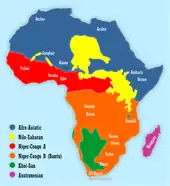
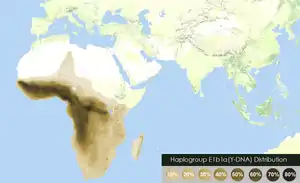
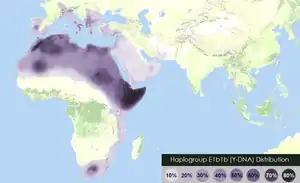

The Father Tongue hypothesis proposes the idea that humans tend to speak their father's language. The hypothesis is based on a 1997 proposal that linguistic affiliation correlates more closely with Y-chromosomal variation than with mitochondrial DNA variation. The initial work was performed on African and European samples by a team of population geneticists, led by Laurent Excoffier. On the basis of these, and similar findings by other geneticists, the hypothesis was elaborated by historical linguist George van Driem in 2010 that the teaching by a mother of her spouse's tongue to her children is a mechanism by which language has preferentially been spread over time. Focusing on prehistoric language shift in already settled areas, examples worldwide[1] show that as little as 10–20% of prehistoric male immigration can (but need not) cause a language switch, indicating an elite imposition such as may have happened with the appearance of the first farmers or metalworkers in the Neolithic, Bronze and Iron Ages.
Early autosomal research
Before the discovery of mtDNA variation and Y-chromosomal variation in the 1980s and 1990s, respectively,[2][3] it was not possible to distinguish male from female effects in population genetics. Instead, researchers had to rely on autosomal variation, starting with the first population genetic study using blood groups by Ludwik Hirszfeld in 1919.[4] Later other genetic polymorphisms were used, for example polymorphisms of proteins of the blood plasma, polymorphisms of human lymphocyte antigens or polymorphisms of immunoglobulins.[5] On this basis, correlations between languages and genetic variation occasionally were proposed,[6][7] but sex-specific questions could not be addressed until the 1990s, when both mtDNA and Y-chromosomal variation in humans became available for study.
Origin of the hypothesis
The Y chromosome follows patrilineal inheritance, meaning it is only passed on among males, from father to son. Mitochondrial DNA on the other hand follows matrilineal inheritance, meaning it is only passed on from the mother to her children and from her daughters to their children. In 1997 Laurent Excoffier, his student Estella Poloni and his team reported that they had found a strong correlation between the Y-chromosomal sequence P49a,f/Taql variation and linguistics, while not being able to find such a correspondence for the mtDNA variation. Poloni et al. proposed the possible consequences of such a correlation, i.e. the Father Tongue hypothesis:
- "As a consequence, the female-specific diversity of our genome would fit less well with geography and linguistics than would our male-specific component. [...] If that were to prove to be the case, then the common belief that we speak our mother's tongue should be revised in favor of the concept of a ‘father's tongue’."[8]
Estella Poloni also presented the Father Tongue hypothesis at an international conference in Paris in April 2000.[9]
On the basis of this population genetic work, historical linguist George van Driem elaborated the Father Tongue hypothesis in his ethnolinguistic publications and in population genetic publications which he has co-authored.[10] At the Indo-Pacific Prehistory Association conference in Taipei in 2002 he proposed that
- "a mother teaching her children their father’s tongue has been a recurrent, ubiquitous and prevalent pattern throughout linguistic history, […] some of the mechanisms of language change over time are likely to be inherent to the dynamics of this pathway of transmission. Such correlations are observed worldwide."[11]
Discovery of Y-chromosomal markers for languages
The next development was the discovery of specific Y-chromosomal markers linked to a language.[12][13][14] These Y-chromosomal variants do not cause language change, but happened to be carried by the historic or prehistoric male speakers spreading the language. These language-specific Y-chromosomal markers create correlations such as those observed by Poloni et al. 1997, and furthermore allow the geographic extent, the time depth and the male immigration level underlying an unrecorded (prehistoric) language change to be determined.
Examples of father tongues
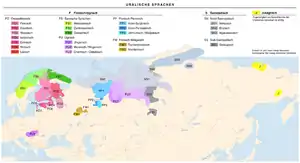
.PNG.webp)
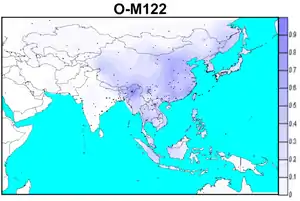
There are several salient examples where the prehistoric diffusion of a language family correlates strongly with the diffusion of Y-chromosomal haplogroups.[1]
- The dispersal of Indo-Europeans from a proposed homeland in the Pontic–Caspian steppe according to the Kurgan hypothesis is suggested to be linked to the spread of the R haplogroup subclade, R1a1, into Europe.[12] R1a1 may also reflect the arrival of the Indo-Aryans into northern India.[10][15]
- The Y-chromosomal lineage L could potentially reflect an earlier patrilingual dispersal of Elamo-Dravidian emanating from a region in modern day Iran.[15] However, the Elamo-Dravidian proposal continues to be rejected by mainstream linguists.
- Austroasiatic speakers show a high frequency of the O2a haplogroup subclade. For example, Munda speakers in north and northeast India show high frequencies of O2a, not found in their regional neighbours who speak languages other than Austroasiatic, whilst their mtDNA haplogroups seem to be those frequent in their region independent of language affinity.[13]
- A population genetic study of 23 Han Chinese populations[16] has shown that the Han expansion southward during the sinification of what today is southern China was predominantly male-biased and is an uncontroversial example of the Father Tongue hypothesis.[10]
- It has also been suggested that Bantu and other Niger-Congo languages correlate well with Y-chromosomal haplogroups.[17][18]
- The spread of Afroasiatic languages has been linked to the expansion of E1b1b haplogroup.[19]
Implications
The Father Tongue hypothesis has far-reaching implications for several processes in linguistics such as language change, language acquisition and sociolinguistics. The Father Tongue hypothesis also has implications for language acquisition, as the hypothesis suggests an evolutionary explanation for why females may be better in some aspects of language performance and acquisition.[20][21][22][23][24][25][26][27][28]
The historical linguist George van Driem interpreted the correlation of Y-chromosomal haplogroups and language families as indicating that the spread of language families was often mediated by male-biased migration, whether these intrusions were martial or something less spectacular. He conjectured that the majority of language communities spoke father tongues rather than mother tongues.[15]
The Father Tongue hypothesis has implications for linguists' understanding of language change. It must be assumed that the dynamics of language change whereby mothers pass on the language of their spouses to their offspring differ from the dynamics of language change in a monolingual community and even from the dynamics of change in a bilingual community where mothers pass on their own language to their children.[29] As a consequence, such dynamics can introduce a discontinuity with the past. For example, it has been observed that Michif, genetically an Algonquian language (like Plains Cree), was relexified by Métis women with Métis French, the language of their husbands, and so the genetic affinity of Michif has come to be almost unidentifiable.[30][31][32] If the process of relexification went beyond the possibility of linguistic reconstruction, the dynamics of such a process may obscure the true linguistic heritance of a community.[29]
Exceptions
Genetics does not determine the language spoken by a human being, and the link between Y-chromosomal haplogroups and linguistic affinities is an observed correlation and not a causal link. While father tongues predominate, exceptions to father tongues exist in the world. Two very well-known exceptions are the Balti in northern Pakistan and Hungarians. The mtDNA haplogroups most frequent among Balti are the same as those of the neighbouring Tibetan communities, whereas the Y-chromosomal haplogroups most frequent in Balti males appear to have entered Baltistan from the west with the introduction of Islam. The Balti speak one of the most conservative Tibetan languages.[10] The language of the Balti corresponds to the mtDNA and not to the Y chromosome and is in effect a salient example of a mother tongue.[15] The other well-known exception is Hungarian . The N1c haplogroup of the Y chromosome, distinguished by Tat-C deletion, is found at a high frequency throughout Uralic language communities, but is virtually missing in Hungarian males. Therefore, while the intrusion of the Magyars into what is today Hungary is historically attested and has left clear linguistic evidence, genetically the Magyar intrusion has left no salient genetic traces. Instead, from a genetic point of view, Hungarians strongly resemble a Western Slavic language community.[29]
See also
References
- 1 2 Forster, Peter; Renfrew C (2011). "Mother Tongue and Y Chromosomes". Science. 333 (6048): 1390–1391. Bibcode:2011Sci...333.1390F. doi:10.1126/science.1205331. PMID 21903800. S2CID 43916070.
- ↑ Ferris, SD; Brown WM, Davidson WS, Wilson AC (1981). "Extensive polymorphism in the mitochondrial DNA of apes". Proceedings of the National Academy of Sciences of the USA. 78 (10): 6319–6323. Bibcode:1981PNAS...78.6319F. doi:10.1073/pnas.78.10.6319. PMC 349030. PMID 6273863.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ↑ Underhill, Peter; Jin L, Lin AA, Mehdi SQ, Jenkins T, Vollrath D, Davis RW, Cavalli-Sforza LL, Oefner PJ (1997). "Detection of numerous Y chromosome biallelic polymorphisms by denaturing high-performance liquid chromatography". Genome Research. 7 (10): 996–1005. doi:10.1101/gr.7.10.996. PMC 310671. PMID 9331370.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ↑ Hirszfeld, L; Hirszfeld H (1919). "Essai d'application des méthodes sérologiques au probléme des races". Anthropologie. 29: 505–537.
- ↑ Cavalli-Sforza, Luigi Luca; Menozzi P, Piazza A (1994). The history and Geography of Human Genes. Princeton, New Jersey: Princeton University Press.
- ↑ Darlington, CD (1947). "The genetic component of language". Heredity. 1 (3): 269–286. doi:10.1038/hdy.1947.18.
- ↑ Cavalli-Sforza, LL; Piazza A, Menozzi P, Mountain J (1988). "Reconstruction of human evolution: bringing together genetic, archaeological, and linguistic data". Proceedings of the National Academy of Sciences of the USA. 85 (16): 6002–6006. Bibcode:1988PNAS...85.6002C. doi:10.1073/pnas.85.16.6002. PMC 281893. PMID 3166138.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ↑ Poloni, Estela S; Semino O, Passarino G, Santachiara-Benerecetti AS, Dupanloup I, Langaney A, Excoffier L (1997). "Human genetic affinities for Y-chromosome p49a,f/TaqI haplotypes show strong correspondence with linguistics". American Journal of Human Genetics. 61 (5): 1015–1035. doi:10.1086/301602. PMC 1716025. PMID 9346874.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ↑ Poloni, Estella Simone; et al. (2000). "Languages and genes: Modes of transmission observed through the analysis of male-specific and female-specific genes". In Jean-Louis Dessalles and Laleh Ghadakpour (ed.). Proceedings: Evolution of Language, 3rd International Conference 3-6 April 2000. Paris: École Nationale Supérieure des Télécommunications. pp. 185–186.
- 1 2 3 4 van Driem, George (2007). "Austroasiatic phylogeny and the Austroasiatic homeland in light of recent population genetic studies". Mon-Khmer Studies. 37: 1–14.
- ↑ van Driem, George (2012). "The ethnolinguistic identity of the domesticators of Asian rice" (PDF). Comptes Rendus Palevol. 11 (2): 117–132. doi:10.1016/j.crpv.2011.07.004.
- 1 2 Zerjal, Tatiana; Pandya A, Santos FR, Adhikari R, Tarazona E, Kayser M, Evgrafov O, Singh L, Thangaraj K, Destro-Bisol G, Thomas MG, Qamar R, Mehdi SQ, Rosser ZH, Hurles ME, Jobling MA, Tyler-Smith C (1999). "The use of Y-chromosomal DNA variation to investigate population history: Recent male spread in Asia and Europe". In Surinder S Papiha, Ranjan Deka and Ranajit Chakraborty (ed.). Genomic Diversity. Applications in Human Population Genetic Studies. Williamsburg, Virginia 26 July - 1 Aug 1998. New York: Kluwer Academic/Plenum Publishers. pp. 91–101.
{{cite book}}: CS1 maint: multiple names: authors list (link) - 1 2 Chaubey, Gyaneshwer; et al. (2010). "Population genetic structure in Indian Austroasiatic speakers: The role of landscape barriers and sex-specific admixture". Molecular Biology and Evolution. 28 (2): 1013–1024. doi:10.1093/molbev/msq288. PMC 3355372. PMID 20978040.
- ↑ Rai, Niraj; et al. (2012). "The phylogeography of Y-chromosome haplogroup H1a1a-M82 reveals the likely Indian origin of the European Romani populations". PLOS ONE. 7 (11): e48477. Bibcode:2012PLoSO...748477R. doi:10.1371/journal.pone.0048477. PMC 3509117. PMID 23209554.
- 1 2 3 4 van Driem, George (2012). "Etyma, shouldered adzes and molecular variants". In Andrea Ender, Adrian Leemann and Bernhard Wälchli (ed.). Methods in Contemporary Linguistics. Berlin: Mouton de Gruyter.
- ↑ Wen, Bo; et al. (2004). "Genetic evidence supports demic diffusion of Han culture". Nature. 431 (7006): 302–305. Bibcode:2004Natur.431..302W. doi:10.1038/nature02878. PMID 15372031. S2CID 4301581.
- ↑ Wood, Elizabeth T; et al. (2005). "Contrasting patterns of Y chromosome and mtDNA variation in Africa: evidence for sex-biased demographic processes". European Journal of Human Genetics. 13 (7): 867–876. doi:10.1038/sj.ejhg.5201408. PMID 15856073.
- ↑ de Filippo, Cesare; et al. (2011). "Y-Chromosomal Variation in Sub-Saharan Africa: Insights Into the History of Niger-Congo Groups". Molecular Biology and Evolution. 28 (3): 1255–1269. doi:10.1093/molbev/msq312. PMC 3561512. PMID 21109585.
- ↑ Gebremeskel, Eyoab I; Ibrahim, Muntaser E (December 2014). "Y-chromosome E haplogroups: their distribution and implication to the origin of Afro-Asiatic languages and pastoralism". European Journal of Human Genetics. 22 (12): 1387–1392. doi:10.1038/ejhg.2014.41. ISSN 1476-5438. PMC 4231410. PMID 24667790.
- ↑ Shaywitz, B. A; et al. (1995). "Sex differences in the functional organization of the brain for language". Nature. 373 (6515): 607–609. Bibcode:1995Natur.373..607S. doi:10.1038/373607a0. PMID 7854416. S2CID 4315684.
- ↑ Pugh, K. R; et al. (1996). "Cerebral organization of component processes in reading". Brain. 119 (4): 1221–1238. doi:10.1093/brain/119.4.1221. PMID 8813285.
- ↑ Pugh, K. R; et al. (1997). "Predicting reading performance from neuroimaging profiles: The cerebral basis of phonological effects in printed word identification". Journal of Experimental Psychology: Human Perception and Performance. 23 (2): 299–318. doi:10.1037/0096-1523.23.2.299. PMID 9103996.
- ↑ Jaeger; et al. (1998). "Sex differences in brain regions activated by grammatical and reading tasks". NeuroReport. 9 (12): 2803–2807. doi:10.1097/00001756-199808240-00022. PMID 9760124. S2CID 41971376.
- ↑ Kansaku, K; et al. (2000). "Sex differences in lateralization revealed in the posterior language areas". Cerebral Cortex. 10 (9): 866–872. doi:10.1093/cercor/10.9.866. PMID 10982747.
- ↑ Rossell, S. L; et al. (2002). "Sex differences in functional brain activation during a lexical visual field task". Brain and Language. 80 (1): 97–105. doi:10.1006/brln.2000.2449. PMID 11817892. S2CID 34669770.
- ↑ Baxter, L. C; et al. (2003). "Sex differences in semantic language processing: A functional MRI study". Brain and Language. 84 (2): 264–272. doi:10.1016/s0093-934x(02)00549-7. PMID 12590915. S2CID 12658733.
- ↑ Clements, A. M; et al. (2006). "Sex differences in cerebral laterality of language and visuospatial processing". Brain and Language. 98 (2): 150–158. doi:10.1016/j.bandl.2006.04.007. PMID 16716389. S2CID 32525196.
- ↑ Burman, Douglas D.; et al. (2008). "Sex differences in neural processing of language among children". Neuropsychologia. 46 (5): 1349–1362. doi:10.1016/j.neuropsychologia.2007.12.021. PMC 2478638. PMID 18262207.
- 1 2 3 van Driem, George (2008). "Reflections on the ethnolinguistic prehistory of the greater Himalayan region". In Brigitte Huber, Marianne Volkart and Paul Widmer (ed.). Chomolangma, Demawend und Kasbek: Festschrift für Roland Bielmeier zu seinem 65. Geburtstag (2 vols.). Halle: International Institute for Tibetan and Buddhist Studies. pp. 39–59.
- ↑ Bakker, Pieter Jan (1992). A language of our own: The genesis of Michif, the mixed Cree-French language of the Canadian Métis. Amsterdam: Universiteit van Amsterdam: doctoral dissertation.
- ↑ Bakker, Pieter Jan (1994). "Michif, the Cree-French mixed language of the Metis buffalo hunters in Canada". In Peter Bakker and Maarten Mous (ed.). Mixed Languages, 15 Case Studies in Language Intertwining (Studies in Language and Language Use, 13). Amsterdam: Instituut voor Fundamenteel Onderzoek naar Taal en Taalgebruik. pp. 13–33.
- ↑ van Driem, George (2001). Languages of the Himalayas. Leiden: Brill.