
Ferlins are an ancient protein family involved in vesicle fusion and membrane trafficking. Ferlins are distinguished by their multiple tandem C2 domains, and sometimes a FerA and a DysF domain. Mutations in ferlins can cause human diseases such as muscular dystrophy and deafness. Abnormalities in expression of myoferlin, a human ferlin protein, is also directly associated with higher mortality rate and tumor recurrence in several types of cancer, including pancreatic, colorectal, breast, cervical, stomach, ovarian, cervical, thyroid, endometrial, and oropharyngeal squamous cell carcinoma.[1][2] In other animals, ferlin mutations can cause infertility.
Ferlins are type II transmembrane proteins (N-terminus on the cytoplasmic side of the membrane) and contain five to seven C2 domains linked in tandem and have a single-pass transmembrane domain located at the C-terminus.[3] The C2 domains are denoted in order from amino-terminus to carboxyl-terminus as C2A to C2G. C2 domains are essentially calcium and phospholipid binding domains, evolved for cell membrane interactions. In fact, many proteins involved in signal transduction, membrane trafficking, and membrane fusion employ C2 domains to target the cell membrane. However, ferlins are unique for containing more C2 domains than any other proteins (between five and seven).[4] FerA and DysF are two intermediate domains that are unique to ferlins. There is less known about FerA and DysF domains, however, mutations of these domains in dysferlin can also lead to muscular dystrophy.[5][6]
As in other mammals, there are six ferlin genes in humans (Fer1L1-Fer1L6). Among them, Fer1L1-Fer1L3 have known disease relevance. Therefore, Fer1L1-Fer1L3 are better characterized compare to Fer1L4-Fer1L6 with unknown function and tissue localization.[7] Fer1L1-Fer1L3 proteins each has a unique name and they correspond to dysferlin, myoferlin, and otoferlin accordingly.[4]
Discovery
The first member of ferlin protein family, fer-1, was discovered in nematode Caenorhabditis elegans. Fer-1 gene was first described in 1997 by Achanzar and Ward.[8] Fer-1 is required for reproduction in C. elegans and was therefore named Fer-1 because of its involvement in fertility. The name is an abbreviation for “fertilization factor 1”. The nomenclature in other ferlins in humans is Fer1Lx, where x is a number from 1-6, each identifying one of the six Fer1-like ferlins in humans.[9]
Evolution
Ferlins are ancient proteins and they have been identified in protists and metazoans, and are known to exist in a range of organisms from unicellular eukaryotes to humans, suggesting primordial functions for ferlins.[7] More specifically, DysF domain and the last two C-terminal C2 domains followed by the C-terminal transmembrane domain (C2E-C2F-TM, containing approximately 489 amino acids) show a high degree of conservation. All ferlins contain several C2 domains. However, C2A may be missing in some ferlins.[7] More specifically, from six human ferlins, three of them do not contain C2A domains. Another highly conserved domain is the N-terminal C2-FerI-C2 sequence. FerI is a motif detected by Pfam, however, the function of this conserved motif is currently unknown.[7][10] Ferlins have been evolved into two groups, DysF-containing and non-DysF ferlins. Most invertebrates possess two ferlin proteins, one from each class. Most vertebrate however, have six ferlin genes, three of which DysF containing and the other three non-DysF ferlins, indicating that vertebrate ferlins are evolved and originated from the two ferlins in early metazoans. Both subgroups have been identified in early metazoans, suggesting the fundamental role associated to these proteins.[7]
Structure
| Dysferlin DysF | |
|---|---|
 Crystal structure of the inner human DysF domain of dysferlin | |
| Identifiers | |
| Symbol | 4CAH |
| SMART | SM00694 |
Ferlins are large proteins and currently the full length structure of ferlins is unknown. In order to understand their structural aspects, ferlin domains have been studied individually:
C2A domains are calcium and lipid binding domains made from 8 β-strands forming 2 sheets. The loops connecting the sheets form the calcium binding site. The β-sheet structure is conserved among C2 domains, however, the loops may have different features.[11] Depending on the amino acids located at the calcium binding site and the loops, C2 domains can have different specificities for calcium and lipid binding, suggesting that they are evolved to function in different environments.[12]
The DysF domain exists as an internal duplication where an inner DysF domain is surrounded by an outer DysF domain. Such structure is a result of gene duplication and both inner and outer DysF domains have adopted the same fold. The structure of DysF is mainly consist of two antiparallel long β-strands. To date, the crystallographic structure of human dysferlin and solution NMR structure of myoferlin DysF have been obtained by Altin Sula et al. and PryankPatel et al. accordingly. Myoferlin and dysferlin DysF domains show 61% sequence identity. A unique feature of DysF domains in both dysferlin and myoferlin is that these domains are held together by arginine/aromatic sidechain (specially tryptophan) stacking.[5][13]
FerA had been predicted using Pfam and SMART and remained uncharacterized both structurally and functionally until recently.[7][14][15] It had been determined by secondary structure prediction however, that FerA domain contains several helices. Recently, a model of FerA structure obtained by homology models have been confirmed by fitting the calculated model into the FerA structure obtained by small-angle X-ray scattering (SAXS) experiments. These structural models provided evidence that FerA contains four helices, which fold to form a four-helix bundle.
Function

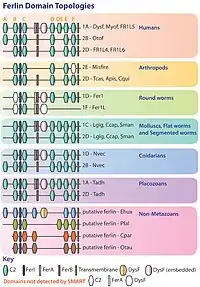
Ferlins play roles in vesicle fusion and membrane trafficking. Different ferlins are found in various organs and they play specific roles.[16] Fer-1 is a member of ferlin protein family, and a fertilization factor involved in fusion of vesicles called membraneous organelles with the sperm plasma membrane during spermatogenesis in C. elegans. In C. elegans spermatids are immobile and during sperm maturation mobility is gained after fusion of membraneous organelles with the plasma membrane. At this point, spermatids extend their pseudopod and become mobile. This process is calcium-dependent and a normal progression of this step requires ferlin's involvement.[17][8][18]
Dysferlin is highly expressed in skeletal muscles, but is also found in heart, placenta, liver, lung, kidney and pancreas. Dysferlin is essential for membrane repair mechanism in muscle cells. Dysferlin in sea stars is 46.9% identical to human dysferlin, and is critical for normal endocytosis during oogenesis and embryogenesis.[19] In humans, dysferlin's primary function is believed to be involvement in muscle membrane repair mechanism. Skeletal muscles experience micro-damages during exercising and daily activities. When muscles are damaged, dysferlin containing vesicles accumulate at the site of injury, and by fusing together and to the membrane, they patch the leakage. In dysferlin-null muscles, these vesicles still accumulate at the damage site, but they cannot fuse and therefore, are unable to repair the damaged muscle cells.[20][21] Otoferlin is another ferlin member in humans and it plays a role in exocytosis of synaptic vesicles at the auditory inner hair cell ribbon synapse.[22] In adult fruit flies, a ferlin member called misfire is expressed in testis and ovaries.[23] Mutations in misfire and Fer-1, ferlins in flies and C. elegans, cause male sterility because of defects in fertilization.[17][23]
Function of ferlin proteins involves employing multiple domains. C2A domains are specialized in lipid binding. The phospholipid interaction is often calcium dependent as C2 domains have evolved to respond to increase in calcium concentration. A sudden increase in calcium concentration is observed in synaptic vesicles or inside muscle cells after membrane damage. Therefore, C2 domains are often referred to as the calcium sensor of C2 domain-containing proteins. The function and mechanism of function of C2 domains is well-characterized, although it may vary between different C2 domains. In general, C2 domains interact with the membrane via electrostatic or hydrophobic interactions.[11] It has been proposed that FerA may be involved in membrane interaction as well. It can in fact interact with neutral or negatively charged phospholipids and the interaction is enhanced in the presence of calcium ions.[6] The molecular mechanism by which FerA interacts with the membrane or calcium ions however, is currently unknown.
Disease association
The most important disease relevance of ferlins in humans is related to mutations in dysferlin. In humans, disease causing mutations in dysferlin have been identified in all C2 domains, FerA domain, DysF domain, and even linker segments.[20] Lack of functional dysferlin causes a group of muscular dystrophies called dysferlinopathies.[24] Dysferlinopathies include limb-girdle muscular dystrophy (LGMD) 2B, Miyoshi myopathy (MM) and distal myopathy of the anterior tibialis.[25] C2A mutations which affect its calcium binding or lipid binding can often cause muscular dystrophy. Interestingly, dysferlin C2B does not bind calcium, however, mutations in this domain can still cause muscular dystrophy.[26] Some mutations in C2A can disrupt dysferlin interaction with other important proteins involved in membrane repair process (such as MG53) which can also lead to muscular dystrophy.[27] Many mutations in dysferlin occur in DysF domain which often disrupt Arginine/Tryptophan stacks of this domain. This leads to a less stable and possibly unfolded protein which may result in the degradation of the entire dysferlin.[13] Several FerA mutations have been also identified. These mutations have been shown to lower the stability of FerA domains which may explain the pathogenicity of these mutations.[6]
Otoferlin has been shown to interact with SNAREs and play a role in a calcium-dependent exocytosis in the hair cells in the inner ear.[28] Mutations in otoferlin can cause mild to profound non-syndromic recessive hearing loss in humans.[16][29][30]
Currently, there is no association between myoferlin mutations and human diseases. However, it has been shown experimentally that loss of myoferlin results in reduced myoblast fusion and muscle size.[31] There is also a correlation between myoferlin overexpression and several types of cancers such as lung cancer and breast cancer.[32][33] In pancreatic ductal adenocarcinoma (PDAC) myoferlin increases cell proliferation and promotes tumorigenesis and its expression negatively correlates with tumor size.[34] Breast cancer patients with overexpressed myoferlin have a lower survival rate. Although it is not yet clear how myoferlin contributes in cancer pathology in a molecular level, there are scientific evidences that myoferlin overexpression is associated with tumor growth and metastasis. In fact, myoferlin depletion in cancer cell lines can result in reduced tumor size and metastasis rate.[35]
References
- ↑ Eisenberg MC, Kim Y, Li R, Ackerman WE, Kniss DA, Friedman A (December 2011). "Mechanistic modeling of the effects of myoferlin on tumor cell invasion". Proceedings of the National Academy of Sciences of the United States of America. 108 (50): 20078–83. Bibcode:2011PNAS..10820078E. doi:10.1073/pnas.1116327108. PMC 3250187. PMID 22135466.
- ↑ Kumar B, Brown NV, Swanson BJ, Schmitt AC, Old M, Ozer E, Agrawal A, Schuller DE, Teknos TN, Kumar P (April 2016). "High expression of myoferlin is associated with poor outcome in oropharyngeal squamous cell carcinoma patients and is inversely associated with HPV-status". Oncotarget. 7 (14): 18665–77. doi:10.18632/oncotarget.7625. PMC 4951318. PMID 26919244.
- ↑ Bansal D, Campbell KP (April 2004). "Dysferlin and the plasma membrane repair in muscular dystrophy". Trends in Cell Biology. 14 (4): 206–13. doi:10.1016/j.tcb.2004.03.001. PMID 15066638.
- 1 2 Coleman BI, Saha S, Sato S, Engelberg K, Ferguson DJ, Coppens I, Lodoen MB, Gubbels MJ (October 2018). "A Member of the Ferlin Calcium Sensor Family Is Essential for Toxoplasma gondii Rhoptry Secretion". mBio. 9 (5). doi:10.1128/mBio.01510-18. PMC 6168857. PMID 30279285.
- 1 2 Patel P, Harris R, Geddes SM, Strehle EM, Watson JD, Bashir R, Bushby K, Driscoll PC, Keep NH (June 2008). "Solution structure of the inner DysF domain of myoferlin and implications for limb girdle muscular dystrophy type 2b" (PDF). Journal of Molecular Biology. 379 (5): 981–90. doi:10.1016/j.jmb.2008.04.046. PMID 18495154.
- 1 2 3 Harsini FM, Chebrolu S, Fuson KL, White MA, Rice AM, Sutton RB (July 2018). "FerA is a Membrane-Associating Four-Helix Bundle Domain in the Ferlin Family of Membrane-Fusion Proteins". Scientific Reports. 8 (1): 10949. Bibcode:2018NatSR...810949H. doi:10.1038/s41598-018-29184-1. PMC 6053371. PMID 30026467.
- 1 2 3 4 5 6 Lek A, Lek M, North KN, Cooper ST (July 2010). "Phylogenetic analysis of ferlin genes reveals ancient eukaryotic origins". BMC Evolutionary Biology. 10: 231. doi:10.1186/1471-2148-10-231. PMC 2923515. PMID 20667140.
- 1 2 Achanzar WE, Ward S (May 1997). "A nematode gene required for sperm vesicle fusion". Journal of Cell Science. 110 (Pt 9) (9): 1073–81. doi:10.1242/jcs.110.9.1073. PMID 9175703.
- ↑ "Which Ferlin protein was discovered first? | Jain Foundation". www.jain-foundation.org. Retrieved 2018-11-18.
- ↑ Staub E, Fiziev P, Rosenthal A, Hinzmann B (May 2004). "Insights into the evolution of the nucleolus by an analysis of its protein domain repertoire". BioEssays. 26 (5): 567–81. doi:10.1002/bies.20032. PMID 15112237.
- 1 2 Rizo J, Südhof TC (June 1998). "C2-domains, structure and function of a universal Ca2+-binding domain". The Journal of Biological Chemistry. 273 (26): 15879–82. doi:10.1074/jbc.273.26.15879. PMID 9632630.
- ↑ Nalefski EA, Wisner MA, Chen JZ, Sprang SR, Fukuda M, Mikoshiba K, Falke JJ (March 2001). "C2 domains from different Ca2+ signaling pathways display functional and mechanistic diversity". Biochemistry. 40 (10): 3089–100. doi:10.1021/bi001968a. PMC 3862187. PMID 11258923.
- 1 2 Sula A, Cole AR, Yeats C, Orengo C, Keep NH (January 2014). "Crystal structures of the human Dysferlin inner DysF domain". BMC Structural Biology. 14: 3. doi:10.1186/1472-6807-14-3. PMC 3898210. PMID 24438169.
- ↑ "SMART: FerA domain annotation". smart.embl-heidelberg.de. Retrieved 2018-11-17.
- ↑ "Family: FerA (PF08165)". Retrieved 2018-11-17.
- 1 2 Lek A, Evesson FJ, Sutton RB, North KN, Cooper ST (February 2012). "Ferlins: regulators of vesicle fusion for auditory neurotransmission, receptor trafficking and membrane repair". Traffic. 13 (2): 185–94. doi:10.1111/j.1600-0854.2011.01267.x. PMID 21838746. S2CID 25445442.
- 1 2 Washington NL, Ward S (June 2006). "FER-1 regulates Ca2+ -mediated membrane fusion during C. elegans spermatogenesis". Journal of Cell Science. 119 (Pt 12): 2552–62. doi:10.1242/jcs.02980. PMID 16735442.
- ↑ Riddle DL, Blumenthal T, Meyer BJ, Priess JR (1997). Organelle Morphogenesis During Spermatogenesis. Cold Spring Harbor Laboratory Press.
- ↑ Oulhen N, Onorato TM, Ramos I, Wessel GM (April 2014). "Dysferlin is essential for endocytosis in the sea star oocyte". Developmental Biology. 388 (1): 94–102. doi:10.1016/j.ydbio.2013.12.018. PMC 3982204. PMID 24368072.
- 1 2 Cooper ST, Head SI (December 2015). "Membrane Injury and Repair in the Muscular Dystrophies". The Neuroscientist. 21 (6): 653–68. doi:10.1177/1073858414558336. PMID 25406223. S2CID 29751207.
- ↑ Demonbreun AR, Quattrocelli M, Barefield DY, Allen MV, Swanson KE, McNally EM (June 2016). "An actin-dependent annexin complex mediates plasma membrane repair in muscle". The Journal of Cell Biology. 213 (6): 705–18. doi:10.1083/jcb.201512022. PMC 4915191. PMID 27298325.
- ↑ Pangršič T, Reisinger E, Moser T (November 2012). "Otoferlin: a multi-C2 domain protein essential for hearing". Trends in Neurosciences. 35 (11): 671–80. doi:10.1016/j.tins.2012.08.002. PMID 22959777. S2CID 12329992.
- 1 2 Smith MK, Wakimoto BT (March 2007). "Complex regulation and multiple developmental functions of misfire, the Drosophila melanogaster ferlin gene". BMC Developmental Biology. 7: 21. doi:10.1186/1471-213X-7-21. PMC 1853072. PMID 17386097.
- ↑ Urtizberea JA, Bassez G, Leturcq F, Nguyen K, Krahn M, Levy N (2008). "Dysferlinopathies". Neurology India. 56 (3): 289–97. doi:10.4103/0028-3886.43447. PMID 18974555.
- ↑ Cacciottolo M, Numitone G, Aurino S, Caserta IR, Fanin M, Politano L, Minetti C, Ricci E, Piluso G, Angelini C, Nigro V (September 2011). "Muscular dystrophy with marked Dysferlin deficiency is consistently caused by primary dysferlin gene mutations". European Journal of Human Genetics. 19 (9): 974–80. doi:10.1038/ejhg.2011.70. PMC 3179367. PMID 21522182.
- ↑ Jin SQ, Yu M, Zhang W, Lyu H, Yuan Y, Wang ZX (October 2016). "Dysferlin Gene Mutation Spectrum in a Large Cohort of Chinese Patients with Dysferlinopathy". Chinese Medical Journal. 129 (19): 2287–93. doi:10.4103/0366-6999.190671. PMC 5040013. PMID 27647186.
- ↑ Matsuda C, Miyake K, Kameyama K, Keduka E, Takeshima H, Imamura T, Araki N, Nishino I, Hayashi Y (November 2012). "The C2A domain in dysferlin is important for association with MG53 (TRIM72)". PLOS Currents. 4: e5035add8caff4. doi:10.1371/5035add8caff4. PMC 3493068. PMID 23145354.
- ↑ Ramakrishnan NA, Drescher MJ, Morley BJ, Kelley PM, Drescher DG (March 2014). "Calcium regulates molecular interactions of otoferlin with soluble NSF attachment protein receptor (SNARE) proteins required for hair cell exocytosis". The Journal of Biological Chemistry. 289 (13): 8750–66. doi:10.1074/jbc.M113.480533. PMC 3979417. PMID 24478316.
- ↑ Choi BY, Ahmed ZM, Riazuddin S, Bhinder MA, Shahzad M, Husnain T, Riazuddin S, Griffith AJ, Friedman TB (March 2009). "Identities and frequencies of mutations of the otoferlin gene (OTOF) causing DFNB9 deafness in Pakistan". Clinical Genetics. 75 (3): 237–43. doi:10.1111/j.1399-0004.2008.01128.x. PMC 3461579. PMID 19250381.
- ↑ Varga R, Avenarius MR, Kelley PM, Keats BJ, Berlin CI, Hood LJ, Morlet TG, Brashears SM, Starr A, Cohn ES, Smith RJ, Kimberling WJ (July 2006). "OTOF mutations revealed by genetic analysis of hearing loss families including a potential temperature sensitive auditory neuropathy allele". Journal of Medical Genetics. 43 (7): 576–81. doi:10.1136/jmg.2005.038612. PMC 2593030. PMID 16371502.
- ↑ Doherty KR, Cave A, Davis DB, Delmonte AJ, Posey A, Earley JU, Hadhazy M, McNally EM (December 2005). "Normal myoblast fusion requires myoferlin". Development. 132 (24): 5565–75. doi:10.1242/dev.02155. PMC 4066872. PMID 16280346.
- ↑ Song DH, Ko GH, Lee JH, Lee JS, Lee GW, Kim HC, Yang JW, Heo RW, Roh GS, Han SY, Kim DC (February 2016). "Myoferlin expression in non-small cell lung cancer: Prognostic role and correlation with VEGFR-2 expression". Oncology Letters. 11 (2): 998–1006. doi:10.3892/ol.2015.3988. PMC 4734036. PMID 26893682.
- ↑ Turtoi A, Blomme A, Bellahcène A, Gilles C, Hennequière V, Peixoto P, Bianchi E, Noel A, De Pauw E, Lifrange E, Delvenne P, Castronovo V (September 2013). "Myoferlin is a key regulator of EGFR activity in breast cancer" (PDF). Cancer Research. 73 (17): 5438–48. doi:10.1158/0008-5472.CAN-13-1142. PMID 23864327.
- ↑ Rademaker G, Hennequière V, Brohée L, Nokin MJ, Lovinfosse P, Durieux F, Gofflot S, Bellier J, Costanza B, Herfs M, Peiffer R, Bettendorff L, Deroanne C, Thiry M, Delvenne P, Hustinx R, Bellahcène A, Castronovo V, Peulen O (August 2018). "Myoferlin controls mitochondrial structure and activity in pancreatic ductal adenocarcinoma, and affects tumor aggressiveness". Oncogene. 37 (32): 4398–4412. doi:10.1038/s41388-018-0287-z. PMC 6085282. PMID 29720728.
- ↑ Li R, Ackerman WE, Mihai C, Volakis LI, Ghadiali S, Kniss DA (2012). "Myoferlin depletion in breast cancer cells promotes mesenchymal to epithelial shape change and stalls invasion". PLOS ONE. 7 (6): e39766. Bibcode:2012PLoSO...739766L. doi:10.1371/journal.pone.0039766. PMC 3384637. PMID 22761893.