| Schizosaccharomyces pombe | |
|---|---|
 | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Fungi |
| Division: | Ascomycota |
| Class: | Schizosaccharomycetes |
| Order: | Schizosaccharomycetales |
| Family: | Schizosaccharomycetaceae |
| Genus: | Schizosaccharomyces |
| Species: | S. pombe |
| Binomial name | |
| Schizosaccharomyces pombe Lindner (1893) | |
| Synonyms[1] | |
| |
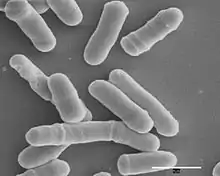
Schizosaccharomyces pombe, also called "fission yeast", is a species of yeast used in traditional brewing and as a model organism in molecular and cell biology. It is a unicellular eukaryote, whose cells are rod-shaped. Cells typically measure 3 to 4 micrometres in diameter and 7 to 14 micrometres in length. Its genome, which is approximately 14.1 million base pairs, is estimated to contain 4,970 protein-coding genes and at least 450 non-coding RNAs.[2]
These cells maintain their shape by growing exclusively through the cell tips and divide by medial fission to produce two daughter cells of equal size, which makes them a powerful tool in cell cycle research.
Fission yeast was isolated in 1893 by Paul Lindner from East African millet beer. The species name pombe is the Swahili word for beer. It was first developed as an experimental model in the 1950s: by Urs Leupold for studying genetics,[3][4] and by Murdoch Mitchison for studying the cell cycle.[5][6][7]
Paul Nurse, a fission yeast researcher, successfully merged the independent schools of fission yeast genetics and cell cycle research. Together with Lee Hartwell and Tim Hunt, Nurse won the 2001 Nobel Prize in Physiology or Medicine for their work on cell cycle regulation.
The sequence of the S. pombe genome was published in 2002, by a consortium led by the Sanger Institute, becoming the sixth model eukaryotic organism whose genome has been fully sequenced. S. pombe researchers are supported by the PomBase MOD (model organism database). This has fully unlocked the power of this organism, with many genes orthologous to human genes identified - 70% to date,[8][9] including many genes involved in human disease.[10] In 2006, sub-cellular localization of almost all the proteins in S. pombe was published using green fluorescent protein as a molecular tag.[11]
Schizosaccharomyces pombe has also become an important organism in studying the cellular responses to DNA damage and the process of DNA replication.
Approximately 160 natural strains of S. pombe have been isolated. These have been collected from a variety of locations including Europe, North and South America, and Asia. The majority of these strains have been collected from cultivated fruits such as apples and grapes, or from the various alcoholic beverages, such as Brazilian Cachaça. S. pombe is also known to be present in fermented tea, kombucha.[12] It is not clear at present whether S. pombe is the major fermenter or a contaminant in such brews. The natural ecology of Schizosaccharomyces yeasts is not well-studied.
History
Schizosaccharomyces pombe was first discovered in 1893 when a group working in a Brewery Association Laboratory in Germany was looking at sediment found in millet beer imported from East Africa that gave it an acidic taste. The term schizo, meaning "split" or "fission", had previously been used to describe other Schizosaccharomycetes. The addition of the word pombe was due to its isolation from East African beer, as pombe means "beer" in Swahili. The standard S. pombe strains were isolated by Urs Leupold in 1946 and 1947 from a culture that he obtained from the yeast collection in Delft, The Netherlands. It was deposited there by A. Osterwalder under the name S. pombe var. liquefaciens, after he isolated it in 1924 from French wine (most probably rancid) at the Federal Experimental Station of Vini- and Horticulture in Wädenswil, Switzerland. The culture used by Urs Leupold contained (besides others) cells with the mating types h90 (strain 968), h- (strain 972), and h+ (strain 975). Subsequent to this, there have been two large efforts to isolate S. pombe from fruit, nectar, or fermentations: one by Florenzano et al.[13] in the vineyards of western Sicily, and the other by Gomes et al. (2002) in four regions of southeast Brazil.[14]
Ecology
The fission yeast S. pombe belongs to the divisio Ascomycota, which represents the largest and most diverse group of fungi. Free-living ascomycetes are commonly found in tree exudates, on plant roots and in surrounding soil, on ripe and rotting fruits, and in association with insect vectors that transport them between substrates. Many of these associations are symbiotic or saprophytic, although numerous ascomycetes (and their basidiomycete cousins) represent important plant pathogens that target myriad plant species, including commercial crops. Among the ascomycetous yeast genera, the fission yeast Schizosaccharomyces is unique because of the deposition of α-(1,3)-glucan or pseudonigeran in the cell wall in addition to the better known β-glucans and the virtual lack of chitin. Species of this genus also differ in mannan composition, which shows terminal d-galactose sugars in the side-chains of their mannans. S. pombe undergo aerobic fermentation in the presence of excess sugar.[15] S. pombe can degrade L-malic acid, one of the dominant organic acids in wine, which makes them diverse among other Saccharomyces strains.
Comparison with budding yeast (Saccharomyces cerevisiae)
The yeast species Schizosaccharomyces pombe and Saccharomyces cerevisiae are both extensively studied; these two species diverged approximately 300 to 600 million years before present,[16] and are significant tools in molecular and cellular biology. Some of the technical discriminants between these two species are:
- S. cerevisiae has approximately 5,600 open reading frames; S. pombe has approximately 5,070 open reading frames.
- Despite similar gene numbers, S. cerevisiae has only about 250 introns, while S. pombe has nearly 5,000.
- S. cerevisiae has 16 chromosomes, S. pombe has 3.
- S. cerevisiae is often diploid while S. pombe is usually haploid.
- S. pombe has a shelterin-like telomere complex while S. cerevisiae does not.[17]
- S. cerevisiae is in the G1 phase of the cell cycle for an extended period (as a consequence, G1-S transition is tightly controlled), while S. pombe remains in the G2 phase of the cell cycle for an extended period (as a consequence, G2-M transition is under tight control).
- Both species share genes with higher eukaryotes that they do not share with each other. S. pombe has RNAi machinery genes like those in vertebrates, while this is missing from S. cerevisiae. S. cerevisiae also has greatly simplified heterochromatin compared to S. pombe.[18] Conversely, S. cerevisiae has well-developed peroxisomes, while S. pombe does not.
- S. cerevisiae has small point centromere of 125 bp, and sequence-defined replication origins of about the same size. On the converse, S. pombe has large, repetitive centromeres (40–100 kb) more similar to mammalian centromeres, and degenerate replication origins of at least 1kb.
S. pombe pathways and cellular processes
S. pombe gene products (proteins and RNAs) participate in many cellular processes common across all life. The fission yeast GO slim provides a categorical high level overview of the biological role of all S. pombe gene products.[8]
Life cycle
The fission yeast is a single-celled fungus with simple, fully characterized genome and a rapid growth rate. It has long been used in brewing, baking, and molecular genetics. S. pombe is a rod-shaped cell, approximately 3 μm in diameter, that grows entirely by elongation at the ends. After mitosis, division occurs by the formation of a septum, or cell plate, that cleaves the cell at its midpoint.
The central events of cell reproduction are chromosome duplication, which takes place in S (Synthetic) phase, followed by chromosome segregation and nuclear division (mitosis) and cell division (cytokinesis), which are collectively called M (Mitotic) phase. G1 is the gap between M and S phases, and G2 is the gap between S and M phases. In the fission yeast, the G2 phase is particularly extended, and cytokinesis (daughter-cell segregation) does not happen until a new S (Synthetic) phase is launched.
Fission yeast governs mitosis by mechanisms that are similar to those in multicellular animals. It normally proliferates in a haploid state. When starved, cells of opposite mating types (P and M) fuse to form a diploid zygote that immediately enters meiosis to generate four haploid spores. When conditions improve, these spores germinate to produce proliferating haploid cells.[19]
 General features of the cell cycle.
General features of the cell cycle. The particular cell cycle of a fission yeast.
The particular cell cycle of a fission yeast.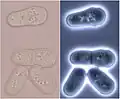 Division stages of Schizosaccharomyces in bright and dark field light microscopy
Division stages of Schizosaccharomyces in bright and dark field light microscopy
Cytokinesis
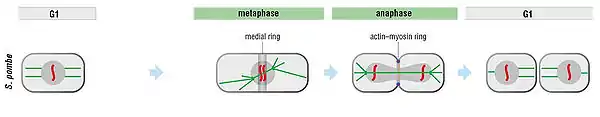
The general features of cytokinesis are shown here. The site of cell division is determined before anaphase. The anaphase spindle (in green on the figure) is then positioned so that the segregated chromosomes are on opposite sides of the predetermined cleavage plane.
Size control
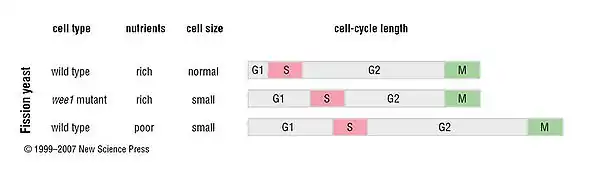
In fission yeast, where growth governs progression through G2/M, a wee1 mutation causes entry into mitosis at an abnormally small size, resulting in a shorter G2. G1 is lengthened, suggesting that progression through Start (beginning of cell cycle) is responsive to growth when the G2/M control is lost. Furthermore, cells in poor nutrient conditions grow slowly and therefore take longer to double in size and divide. Low nutrient levels also reset the growth threshold so that cell progresses through the cell cycle at a smaller size. Upon exposure to stressful conditions [heat (40 °C) or the oxidizing agent hydrogen peroxide] S. pombe cells undergo aging as measured by increased cell division time and increased probability of cell death.[20] Finally, wee1 mutant fission yeast cells are smaller than wild-type cells, but take just as long to go through the cell cycle. This is possible because small yeast cells grow slower, that is, their added total mass per unit time is smaller than that of normal cells.
A spatial gradient is thought to coordinate cell size and mitotic entry in fission yeast.[21][22][23] The Pom1 protein kinase (green) is localized to the cell cortex, with the highest concentration at the cell tips. The cell-cycle regulators Cdr2, Cdr1 and Wee1 are present in cortical nodes in the middle of the cell (blue and red dots). a, In small cells, the Pom1 gradient reaches most of the cortical nodes (blue dots). Pom1 inhibits Cdr2, preventing Cdr2 and Cdr1 from inhibiting Wee1, and allowing Wee1 to phosphorylate Cdk1, thus inactivating cyclin-dependent kinase (CDK) activity and preventing entry into mitosis. b, In long cells, the Pom1 gradient does not reach the cortical nodes (red dots), and therefore Cdr2 and Cdr1 remain active in the nodes. Cdr2 and Cdr1 inhibit Wee1, preventing phosphorylation of Cdk1 and thereby leading to activation of CDK and mitotic entry. (This simplified diagram omits several other regulators of CDK activity.)
Mating-type switching
Fission yeast switches mating type by a replication-coupled recombination event, which takes place during S phase of the cell cycle. Fission yeast uses intrinsic asymmetry of the DNA replication process to switch the mating type; it was the first system where the direction of replication was shown to be required for the change of the cell type. Studies of the mating-type switching system lead to a discovery and characterization of a site-specific replication termination site RTS1, a site-specific replication pause site MPS1, and a novel type of chromosomal imprint, marking one of the sister chromatids at the mating-type locus mat1. In addition, work on the silenced donor region has led to great advances in understanding formation and maintenance of heterochromatin.[24]
Responses to DNA damage
Schizosaccharomyces pombe is a facultative sexual microorganism that can undergo mating when nutrients are limiting.[25] Exposure of S. pombe to hydrogen peroxide, an agent that causes oxidative stress leading to oxidative DNA damage, strongly induces mating and formation of meiotic spores.[26] This finding suggests that meiosis, and particularly meiotic recombination, may be an adaptation for repairing DNA damage.[26] Supporting this view is the finding that single base lesions of the type dU:dG in the DNA of S. pombe stimulate meiotic recombination.[27] This recombination requires uracil-DNA glycosylase, an enzyme that removes uracil from the DNA backbone and initiates base excision repair. On the basis of this finding, it was proposed that base excision repair of either a uracil base, an abasic site, or a single-strand nick is sufficient to initiate recombination in S. pombe.[27] Other experiments with S. pombe indicated that faulty processing of DNA replication intermediates, i.e. Okazaki fragments, causes DNA damages such as single-strand nicks or gaps, and that these stimulate meiotic recombination.[28]
As a model system
Fission yeast has become a notable model system to study basic principles of a cell that can be used to understand more complex organisms like mammals and in particular humans.[29][30] This single cell eukaryote is nonpathogenic and easily grown and manipulated in the lab.[31][32] Fission yeast contains one of the smallest numbers of genes of a known genome sequence for a eukaryote, and has only three chromosomes in its genome.[33] Many of the genes responsible for cell division and cellular organization in fission yeast cell are also found in the human's genome.[31][32][34] Cell cycle regulation and division are crucial for growth and development of any cell. Fission yeast's conserved genes has been heavily studied and the reason for many recent biomedical developments.[35][36] Fission yeast is also a practical model system to observe cell division because fission yeast's are cylindrically shaped single celled eukaryotes that divide and reproduce by medial fission.[31] This can easily be seen using microscopy. Fission yeast also have an extremely short generation time, 2 to 4 hours, which also makes it an easy model system to observe and grow in the laboratory[32] Fission yeast's simplicity in genomic structure yet similarities with mammalian genome, ease of ability to manipulate, and ability to be used for drug analysis is why fission yeast is making many contributions to biomedicine and cellular biology research, and a model system for genetic analysis.[32][25][30][37][38]
Genome
Schizosaccharomyces pombe is often used to study cell division and growth because of conserved genomic regions also seen in humans including: heterochromatin proteins, large origins of replication, large centromeres, conserved cellular checkpoints, telomere function, gene splicing, and many other cellular processes.[33][39][40] S. pombe's genome was fully sequenced in 2002, the sixth eukaryotic genome to be sequenced as part of the Genome Project. An estimated 4,979 genes were discovered within three chromosomes containing about 14Mb of DNA. This DNA is contained within 3 different chromosomes in the nucleus with gaps in the centromeric (40kb) and telomeric (260kb) regions.[33] After the initial sequencing of the fission yeast's genome, other previous non-sequenced regions of the genes have been sequenced. Structural and functional analysis of these gene regions can be found on large scale fission yeast databases such as PomBase.
Forty-three percent of the genes in the Genome Project were found to contain introns in 4,739 genes. Fission yeast does not have as many duplicated genes compared to budding yeast, only containing 5%, making fission yeast a great model genome to observe and gives researchers the ability to create more functional research approaches. S. pombe's having a large number of introns gives opportunities for an increase of range of protein types produced from alternative splicing and genes that code for comparable genes in human.[33] 81% of the three centromeres in fission yeast have been sequenced. The lengths of the three centromeres were found to be 34, 65, and 110 kb. This is 300–100 times longer than the centromeres of budding yeast. An extremely high level of conservation (97%) is also seen over 1,780-bp region in the DGS regions of the centromere. This elongation of centromeres and its conservative sequences makes fission yeast a practical model system to use to observe cell division and in humans because of their likeness.[33][41][42]
PomBase[8][43] reports over 69% of protein coding genes have human orthologs and over 500 of these are associated with human disease . This makes S. pombe a great system to use to study human genes and disease pathways, especially cell cycle and DNA checkpoint systems.[42][44][45][46]
The genome of S. pombe contains meiotic drivers and drive suppressors called wtf genes.[47]
Genetic diversity
Biodiversity and evolutionary study of fission yeast was carried out on 161 strains of Schizosaccharomyces pombe collected from 20 countries.[48] Modeling of the evolutionary rate showed that all strains derived from a common ancestor that has lived since ~2,300 years ago. The study also identified a set of 57 strains of fission yeast that each differed by ≥1,900 SNPs,[48] and all detected 57 strains of fission yeast were prototrophic (able to grow on the same minimal medium as the reference strain).[48] A number of studies on S.pombe genome support the idea that the genetic diversity of fission yeast strains is slightly less than budding yeast.[48] Indeed, only limited variations of S.pombe occur in proliferation in different environments. In addition, the amount of phenotypic variation segregating in fission yeast is less than that seen, in S. cerevisiae.[49] Since most strains of fission yeast were isolated from brewed beverages, there is no ecological or historical context to this dispersal.
Cell cycle analysis
DNA replication in yeast has been increasingly studied by many researchers. Further understanding of DNA replication, gene expression, and conserved mechanisms in yeast can provide researchers with information on how these systems operate in mammalian cells in general and human cells in particular.[40][50][51][52] Other stages, such as cellular growth and aging, are also observed in yeast in order to understand these mechanisms in more complex systems.[34][53][54][55]
S. pombe stationary phase cells undergo chronological aging due to production of reactive oxygen species that cause DNA damages. Most such damages can ordinarily be repaired by DNA base excision repair and nucleotide excision repair.[56] Defects in these repair processes lead to reduced survival.
Cytokinesis is one of the components of cell division that is often observed in fission yeast. Well-conserved components of cytokinesis are observed in fission yeast and allow us to look at various genomic scenarios and pinpoint mutations.[45][57][58] Cytokinesis is a permanent step and very crucial to the wellbeing of the cell.[59] Contractile ring formation in particular is heavily studied by researchers using S. pombe as a model system. The contractile ring is highly conserved in both fission yeast and human cytokinesis.[45] Mutations in cytokinesis can result in many malfunctions of the cell including cell death and development of cancerous cells.[45] This is a complex process in human cell division, but in S. pombe simpler experiments can yield results that can then be applied for research in higher-order model systems such as humans.
One of the safety precautions that the cell takes to ensure precise cell division occurs is the cell-cycle checkpoint.[60][61] These checkpoints ensure any mutagens are eliminated.[62] This is done often by relay signals that stimulate ubiquitination of the targets and delay cytokinesis.[33] Without mitotic check points such as these, mutagens are created and replicated, resulting in multitudes of cellular issues including cell death or tumorigenesis seen in cancerous cells. Paul Nurse, Leland Hartwell, and Tim Hunt were awarded the Nobel Prize in Physiology or Medicine in 2001. They discovered key conserved checkpoints that are crucial for a cell to divide properly. These findings have been linked to cancer and diseased cells and are a notable finding for biomedicine.[63]
Researchers using fission yeast as a model system also look at organelle dynamics and responses and the possible correlations between yeast cells and mammalian cells.[64][65] Mitochondria diseases, and various organelle systems such as the Golgi apparatus and endoplasmic reticulum, can be further understood, by observing fission yeast's chromosome dynamics and protein expression levels and regulation.[46][51][66][67][68][69]
Biomedical tool
However, there are limitations with using fission yeast as a model system: its multidrug resistance. "The MDR response involves overexpression of two types of drug efflux pumps, the ATP-binding cassette (ABC) family... and the major facilitator superfamily".[35] Paul Nurse and some of his colleagues have recently created S. pombe strains sensitive to chemical inhibitors and common probes to see whether it is possible to use fission yeast as a model system of chemical drug research.[35]
For example, Doxorubicin, a very common chemotherapeutic antibiotic, has many adverse side-effects. Researchers are looking for ways to further understand how doxorubicin works by observing the genes linked to resistance by using fission yeast as a model system. Links between doxorubicin adverse side-effects and chromosome metabolism and membrane transport were seen. Metabolic models for drug targeting are now being used in biotechnology, and further advances are expected in the future using the fission yeast model system.[36]
Experimental approaches
Fission yeast is easily accessible, easily grown and manipulated to make mutants, and able to be maintained at either a haploid or diploid state. S. pombe is normally a haploid cell but, when put under stressful conditions, usually nitrogen deficiency, two cells will conjugate to form a diploid that later form four spores within a tetrad ascus.[32] This process is easily visible and observable under any microscope and allows us to look at meiosis in a simpler model system to see how this phenomenon operates.
Virtually any genetics experiment or technique can, therefore, be applied to this model system such as: tetrad dissection, mutagens analysis, transformations, and microscopy techniques such as FRAP and FRET. New models, such as Tug-Of-War (gTOW), are also being used to analyze yeast robustness and observe gene expression. Making knock-in and knock-out genes is fairly easy and with the fission yeast's genome being sequenced this task is very accessible and well known.[70][71]
See also
References
- ↑ "Schizosaccharomyces pombe". Global Biodiversity Information Facility. Retrieved 14 August 2021.
- ↑ Wilhelm BT, Marguerat S, Watt S, Schubert F, Wood V, Goodhead I, et al. (June 2008). "Dynamic repertoire of a eukaryotic transcriptome surveyed at single-nucleotide resolution". Nature. 453 (7199): 1239–43. Bibcode:2008Natur.453.1239W. doi:10.1038/nature07002. PMID 18488015. S2CID 205213499.
- ↑ Leupold U (1950). "Die Vererbung von Homothallie und Heterothallie bei Schizosaccharomyces pombe". C R Trav Lab Carlsberg Ser Physiol. 24: 381–480.
- ↑ Leupold U. (1993) The origins of Schizosaccharomyces pombe genetics. In: Hall MN, Linder P. eds. The Early Days of Yeast Genetics. New York. Cold Spring Harbor Laboratory Press. p 125–128.
- ↑ Mitchison JM (October 1957). "The growth of single cells. I. Schizosaccharomyces pombe". Experimental Cell Research. 13 (2): 244–62. doi:10.1016/0014-4827(57)90005-8. PMID 13480293.
- ↑ Mitchison JM (April 1990). "The fission yeast, Schizosaccharomyces pombe". BioEssays. 12 (4): 189–91. doi:10.1002/bies.950120409. PMID 2185750.
- ↑ Fantes PA, Hoffman CS (June 2016). "A Brief History of Schizosaccharomyces pombe Research: A Perspective Over the Past 70 Years". Genetics. 203 (2): 621–9. doi:10.1534/genetics.116.189407. PMC 4896181. PMID 27270696.
- 1 2 3 Wood V, Harris MA, McDowall MD, Rutherford K, Vaughan BW, Staines DM, et al. (January 2012). "PomBase: a comprehensive online resource for fission yeast". Nucleic Acids Research. 40 (Database issue): D695-9. doi:10.1093/nar/gkr853. PMC 3245111. PMID 22039153.
- ↑ "PomBase".
- ↑ "PomBase".
- ↑ Matsuyama A, Arai R, Yashiroda Y, Shirai A, Kamata A, Sekido S, et al. (July 2006). "ORFeome cloning and global analysis of protein localization in the fission yeast Schizosaccharomyces pombe". Nature Biotechnology. 24 (7): 841–7. doi:10.1038/nbt1222. PMID 16823372. S2CID 10397608.
- ↑ Teoh AL, Heard G, Cox J (September 2004). "Yeast ecology of Kombucha fermentation". International Journal of Food Microbiology. 95 (2): 119–26. doi:10.1016/j.ijfoodmicro.2003.12.020. PMID 15282124.
- ↑ Florenzano G, Balloni W, Materassi R (1977). "Contributo alla ecologia dei lieviti Schizosaccharomyces sulle uve". Vitis. 16: 38–44.
- ↑ Gómez EB, Bailis JM, Forsburg SL (2002). "Fission yeast enters a joyful new era". Genome Biology. 3 (6): REPORTS4017. doi:10.1186/gb-2002-3-6-reports4017. PMC 139370. PMID 12093372.
- ↑ Lin Z, Li WH (April 2011). "The evolution of aerobic fermentation in Schizosaccharomyces pombe was associated with regulatory reprogramming but not nucleosome reorganization". Molecular Biology and Evolution. 28 (4): 1407–13. doi:10.1093/molbev/msq324. PMC 3058771. PMID 21127171.
- ↑ Douzery EJ, Snell EA, Bapteste E, Delsuc F, Philippe H (October 2004). "The timing of eukaryotic evolution: does a relaxed molecular clock reconcile proteins and fossils?". Proceedings of the National Academy of Sciences of the United States of America. 101 (43): 15386–91. Bibcode:2004PNAS..10115386D. doi:10.1073/pnas.0403984101. PMC 524432. PMID 15494441.
- ↑ Price CM, Boltz KA, Chaiken MF, Stewart JA, Beilstein MA, Shippen DE (August 2010). "Evolution of CST function in telomere maintenance". Cell Cycle. 9 (16): 3157–65. doi:10.4161/cc.9.16.12547. PMC 3041159. PMID 20697207.
- ↑ Grunstein, Michael, and Susan Gasser. "Epigenetics in Saccharomyces cerevisiae." Epigenetics. 1. Cold Spring Harbor Press, 2007.
- ↑ Morgan, David O. (2007). The Cell Cycle Principles of Control. London: New Science Press. ISBN 978-0-19-920610-0. OCLC 70173205.
- ↑ Coelho M, Dereli A, Haese A, Kühn S, Malinovska L, DeSantis ME, et al. (October 2013). "Fission yeast does not age under favorable conditions, but does so after stress". Current Biology. 23 (19): 1844–52. doi:10.1016/j.cub.2013.07.084. PMC 4620659. PMID 24035542.
- ↑ Moseley, James B.; Mayeux, Adeline; Paoletti, Anne; Nurse, Paul (2009). "A spatial gradient coordinates cell size and mitotic entry in fission yeast". Nature. 459 (7248): 857–860. Bibcode:2009Natur.459..857M. doi:10.1038/nature08074. ISSN 1476-4687. PMID 19474789. S2CID 4330336.
- ↑ Martin, Sophie G.; Berthelot-Grosjean, Martine (2009-06-11). "Polar gradients of the DYRK-family kinase Pom1 couple cell length with the cell cycle". Nature. 459 (7248): 852–856. Bibcode:2009Natur.459..852M. doi:10.1038/nature08054. ISSN 1476-4687. PMID 19474792. S2CID 4412402.
- ↑ Sawin KE (June 2009). "Cell cycle: Cell division brought down to size". Nature. 459 (7248): 782–3. Bibcode:2009Natur.459..782S. doi:10.1038/459782a. PMID 19516326. S2CID 4402226.
- ↑ Klar, Amar J.S. (2007-12-01). "Lessons Learned from Studies of Fission Yeast Mating-Type Switching and Silencing". Annual Review of Genetics. 41 (1): 213–236. doi:10.1146/annurev.genet.39.073103.094316. ISSN 0066-4197. PMID 17614787.
- 1 2 Davey J (December 1998). "Fusion of a fission yeast". Yeast. 14 (16): 1529–66. doi:10.1002/(SICI)1097-0061(199812)14:16<1529::AID-YEA357>3.0.CO;2-0. PMID 9885154. S2CID 44652765.
- 1 2 Bernstein C, Johns V (April 1989). "Sexual reproduction as a response to H2O2 damage in Schizosaccharomyces pombe". Journal of Bacteriology. 171 (4): 1893–7. doi:10.1128/jb.171.4.1893-1897.1989. PMC 209837. PMID 2703462.
- 1 2 Pauklin S, Burkert JS, Martin J, Osman F, Weller S, Boulton SJ, et al. (May 2009). "Alternative induction of meiotic recombination from single-base lesions of DNA deaminases". Genetics. 182 (1): 41–54. doi:10.1534/genetics.109.101683. PMC 2674839. PMID 19237686.
- ↑ Farah JA, Cromie G, Davis L, Steiner WW, Smith GR (December 2005). "Activation of an alternative, rec12 (spo11)-independent pathway of fission yeast meiotic recombination in the absence of a DNA flap endonuclease". Genetics. 171 (4): 1499–511. doi:10.1534/genetics.105.046821. PMC 1456079. PMID 16118186.
- ↑ Forsburg SL (June 2005). "The yeasts Saccharomyces cerevisiae and Schizosaccharomyces pombe: models for cell biology research". Gravitational and Space Biology Bulletin. 18 (2): 3–9. PMID 16038088.
- 1 2 Forsburg SL, Rhind N (February 2006). "Basic methods for fission yeast". Yeast. 23 (3): 173–83. doi:10.1002/yea.1347. PMC 5074380. PMID 16498704.
- 1 2 3 Wixon J (2002). "Featured organism: Schizosaccharomyces pombe, the fission yeast". Comparative and Functional Genomics. 3 (2): 194–204. doi:10.1002/cfg.92. PMC 2447254. PMID 18628834.
- 1 2 3 4 5 Forsburg SL. "PombeNet". Retrieved 7 February 2016.
- 1 2 3 4 5 6 Wood V, Gwilliam R, Rajandream MA, Lyne M, Lyne R, Stewart A, et al. (February 2002). "The genome sequence of Schizosaccharomyces pombe". Nature. 415 (6874): 871–80. Bibcode:2002Natur.415..871W. doi:10.1038/nature724. PMID 11859360.
- 1 2 Das M, Wiley DJ, Medina S, Vincent HA, Larrea M, Oriolo A, Verde F (June 2007). "Regulation of cell diameter, For3p localization, and cell symmetry by fission yeast Rho-GAP Rga4p". Molecular Biology of the Cell. 18 (6): 2090–101. doi:10.1091/mbc.E06-09-0883. PMC 1877093. PMID 17377067.
- 1 2 3 Kawashima SA, Takemoto A, Nurse P, Kapoor TM (July 2012). "Analyzing fission yeast multidrug resistance mechanisms to develop a genetically tractable model system for chemical biology". Chemistry & Biology. 19 (7): 893–901. doi:10.1016/j.chembiol.2012.06.008. PMC 3589755. PMID 22840777.
- 1 2 Tay Z, Eng RJ, Sajiki K, Lim KK, Tang MY, Yanagida M, Chen ES (24 January 2013). "Cellular robustness conferred by genetic crosstalk underlies resistance against chemotherapeutic drug doxorubicin in fission yeast". PLOS ONE. 8 (1): e55041. Bibcode:2013PLoSO...855041T. doi:10.1371/journal.pone.0055041. PMC 3554685. PMID 23365689.
- ↑ Forsburg SL (September 1999). "The best yeast?". Trends in Genetics. 15 (9): 340–4. doi:10.1016/s0168-9525(99)01798-9. PMID 10461200.
- ↑ Hoffman CS, Wood V, Fantes PA (October 2015). "An Ancient Yeast for Young Geneticists: A Primer on the Schizosaccharomyces pombe Model System". Genetics. 201 (2): 403–23. doi:10.1534/genetics.115.181503. PMC 4596657. PMID 26447128.
- ↑ Sabatinos SA, Mastro TL, Green MD, Forsburg SL (January 2013). "A mammalian-like DNA damage response of fission yeast to nucleoside analogs". Genetics. 193 (1): 143–57. doi:10.1534/genetics.112.145730. PMC 3527242. PMID 23150603.
- 1 2 Hayano M, Kanoh Y, Matsumoto S, Renard-Guillet C, Shirahige K, Masai H (January 2012). "Rif1 is a global regulator of timing of replication origin firing in fission yeast". Genes & Development. 26 (2): 137–50. doi:10.1101/gad.178491.111. PMC 3273838. PMID 22279046.
- ↑ Burrack LS, Berman J (July 2012). "Neocentromeres and epigenetically inherited features of centromeres". Chromosome Research. 20 (5): 607–19. doi:10.1007/s10577-012-9296-x. PMC 3409321. PMID 22723125.
- 1 2 Stimpson KM, Matheny JE, Sullivan BA (July 2012). "Dicentric chromosomes: unique models to study centromere function and inactivation". Chromosome Research. 20 (5): 595–605. doi:10.1007/s10577-012-9302-3. PMC 3557915. PMID 22801777.
- ↑ McDowall MD, Harris MA, Lock A, Rutherford K, Staines DM, Bähler J, et al. (January 2015). "PomBase 2015: updates to the fission yeast database". Nucleic Acids Research. 43 (Database issue): D656-61. doi:10.1093/nar/gku1040. PMC 4383888. PMID 25361970.
- ↑ Kadura S, Sazer S (July 2005). "SAC-ing mitotic errors: how the spindle assembly checkpoint (SAC) plays defense against chromosome mis-segregation". Cell Motility and the Cytoskeleton. 61 (3): 145–60. doi:10.1002/cm.20072. PMID 15887295.
- 1 2 3 4 Lee IJ, Coffman VC, Wu JQ (October 2012). "Contractile-ring assembly in fission yeast cytokinesis: Recent advances and new perspectives". Cytoskeleton. 69 (10): 751–63. doi:10.1002/cm.21052. PMC 5322539. PMID 22887981.
- 1 2 Rinaldi T, Dallabona C, Ferrero I, Frontali L, Bolotin-Fukuhara M (December 2010). "Mitochondrial diseases and the role of the yeast models". FEMS Yeast Research. 10 (8): 1006–22. doi:10.1111/j.1567-1364.2010.00685.x. PMID 20946356.
- ↑ Núñez MAB, Sabbarini IM, Eickbush MT, Liang Y, Lange JJ, Kent AM, Zanders SE (February 2020). "Dramatically diverse Schizosaccharomyces pombe wtf meiotic drivers all display high gamete-killing efficiency". PLOS Genetics. 16 (2): e1008350. doi:10.1371/journal.pgen.1008350. PMC 7032740. PMID 32032353.
- 1 2 3 4 Jeffares, Daniel C.; et al. (2015). "The genomic and phenotypic diversity of Schizosaccharomyces pombe". Nature Genetics. 47 (3): 235–241. doi:10.1038/ng.3215. PMC 4645456. PMID 25665008.
- ↑ Brown, William R. A.; Liti, Gianni; Rosa, Carlos; James, Steve; Roberts, Ian; Robert, Vincent; Jolly, Neil; Tang, Wen; Baumann, Peter; Green, Carter; Schlegel, Kristina; Young, Jonathan; Hirchaud, Fabienne; Leek, Spencer; Thomas, Geraint; Blomberg, Anders; Warringer, Jonas (2011). "A Geographically Diverse Collection of Schizosaccharomyces pombe Isolates Shows Limited Phenotypic Variation but Extensive Karyotypic Diversity". G3: Genes, Genomes, Genetics. 1 (7): 615–626. doi:10.1534/g3.111.001123. PMC 3276172. PMID 22384373.
- ↑ Mojardín L, Vázquez E, Antequera F (November 2013). "Specification of DNA replication origins and genomic base composition in fission yeasts". Journal of Molecular Biology. 425 (23): 4706–13. doi:10.1016/j.jmb.2013.09.023. hdl:10261/104754. PMID 24095860.
- 1 2 Forsburg SL (April 2002). "Only connect: linking meiotic DNA replication to chromosome dynamics". Molecular Cell. 9 (4): 703–11. doi:10.1016/S1097-2765(02)00508-7. PMID 11983163.
- ↑ Moriya H, Chino A, Kapuy O, Csikász-Nagy A, Novák B (December 2011). "Overexpression limits of fission yeast cell-cycle regulators in vivo and in silico". Molecular Systems Biology. 7 (1): 556. doi:10.1038/msb.2011.91. PMC 3737731. PMID 22146300.
- ↑ Das M, Wiley DJ, Chen X, Shah K, Verde F (August 2009). "The conserved NDR kinase Orb6 controls polarized cell growth by spatial regulation of the small GTPase Cdc42". Current Biology. 19 (15): 1314–9. doi:10.1016/j.cub.2009.06.057. PMID 19646873. S2CID 12744756.
- ↑ Moseley JB (October 2013). "Cellular aging: symmetry evades senescence". Current Biology. 23 (19): R871-3. doi:10.1016/j.cub.2013.08.013. PMC 4276399. PMID 24112980.
- ↑ Cooper S (2013). "Schizosaccharomyces pombe grows exponentially during the division cycle with no rate change points" (PDF). FEMS Yeast Res. 13 (7): 650–8. doi:10.1111/1567-1364.12072. PMID 23981297.
- ↑ Senoo T, Kawano S, Ikeda S (March 2017). "DNA base excision repair and nucleotide excision repair synergistically contribute to survival of stationary-phase cells of the fission yeast Schizosaccharomyces pombe". Cell Biology International. 41 (3): 276–286. doi:10.1002/cbin.10722. PMID 28032397. S2CID 39454427.
- ↑ Cadou A, Couturier A, Le Goff C, Xie L, Paulson JR, Le Goff X (March 2013). "The Kin1 kinase and the calcineurin phosphatase cooperate to link actin ring assembly and septum synthesis in fission yeast". Biology of the Cell. 105 (3): 129–48. doi:10.1111/boc.201200042. PMID 23294323. S2CID 21404821.
- ↑ Balazs A, Batta G, Miklos I, Acs-Szabo L, Vazquez de Aldana CR, Sipiczki M (March 2012). "Conserved regulators of the cell separation process in Schizosaccharomyces". Fungal Genetics and Biology. 49 (3): 235–49. doi:10.1016/j.fgb.2012.01.003. hdl:10261/51389. PMID 22300943.
- ↑ Rincon SA, Paoletti A (October 2012). "Mid1/anillin and the spatial regulation of cytokinesis in fission yeast". Cytoskeleton. 69 (10): 764–77. doi:10.1002/cm.21056. PMID 22888038. S2CID 22906028.
- ↑ Das M, Chiron S, Verde F (2010). "Microtubule-dependent spatial organization of mitochondria in fission yeast". Microtubules: In vivo. Methods in Cell Biology. Vol. 97. pp. 203–21. doi:10.1016/S0091-679X(10)97012-X. ISBN 9780123813497. PMID 20719273.
- ↑ Fraser HB (2013). "Cell-cycle regulated transcription associates with DNA replication timing in yeast and human". Genome Biology. 14 (10): R111. arXiv:1308.1985. doi:10.1186/gb-2013-14-10-r111. PMC 3983658. PMID 24098959.
- ↑ Li PC, Green MD, Forsburg SL (2013). "Mutations disrupting histone methylation have different effects on replication timing in S. pombe centromere". PLOS ONE. 8 (5): e61464. Bibcode:2013PLoSO...861464L. doi:10.1371/journal.pone.0061464. PMC 3641051. PMID 23658693.
- ↑ "Sir Paul Nurse - Biographical". The Official Site of the Nobel Prize. 2001. Retrieved 7 February 2016.
- ↑ Zhao J, Lendahl U, Nistér M (March 2013). "Regulation of mitochondrial dynamics: convergences and divergences between yeast and vertebrates". Cellular and Molecular Life Sciences. 70 (6): 951–76. doi:10.1007/s00018-012-1066-6. PMC 3578726. PMID 22806564.
- ↑ Abelovska L (2011). "Mitochondria as protean organelles: membrane processes that influence mitochondrial shape in yeast". General Physiology and Biophysics. 30 Spec No (5): S13-24. doi:10.4149/gpb_2011_SI1_13. PMID 21869447.
- ↑ Chino A, Makanae K, Moriya H (3 September 2013). "Relationships between cell cycle regulator gene copy numbers and protein expression levels in Schizosaccharomyces pombe". PLOS ONE. 8 (9): e73319. Bibcode:2013PLoSO...873319C. doi:10.1371/journal.pone.0073319. PMC 3760898. PMID 24019917.
- ↑ Raychaudhuri S, Young BP, Espenshade PJ, Loewen C (August 2012). "Regulation of lipid metabolism: a tale of two yeasts". Current Opinion in Cell Biology. 24 (4): 502–8. doi:10.1016/j.ceb.2012.05.006. PMC 4339016. PMID 22694927.
- ↑ Babu M, Vlasblom J, Pu S, Guo X, Graham C, Bean BD, et al. (September 2012). "Interaction landscape of membrane-protein complexes in Saccharomyces cerevisiae". Nature. 489 (7417): 585–9. Bibcode:2012Natur.489..585B. doi:10.1038/nature11354. PMID 22940862. S2CID 4344457.
- ↑ Suda Y, Nakano A (April 2012). "The yeast Golgi apparatus". Traffic. 13 (4): 505–10. doi:10.1111/j.1600-0854.2011.01316.x. PMID 22132734.
- ↑ "Trans-NIH.pombe Initiative". 2002.
{{cite journal}}: Cite journal requires|journal=(help) - ↑ Green, M. D. Sabatinos, S. A. Forsburg, S. L. (2009). "Microscopy Techniques to Examine DNA Replication in Fission Yeast". DNA Replication. Methods in Molecular Biology. Vol. 521. pp. 463–82. doi:10.1007/978-1-60327-815-7_26. ISBN 978-1-60327-814-0. PMID 19563123.
{{cite book}}: CS1 maint: multiple names: authors list (link)