| Histamine N-methyltransferase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| EC no. | 2.1.1.8 | ||||||||
| CAS no. | 9029-80-5 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| Gene Ontology | AmiGO / QuickGO | ||||||||
| |||||||||
| histamine N-methyltransferase | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Identifiers | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Aliases | HNMT, HMT, HNMT-S1, HNMT-S2, MRT51 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| External IDs | OMIM: 605238 MGI: 2153181 HomoloGene: 5032 GeneCards: HNMT | ||||||||||||||||||||||||||||||||||||||||||||||||||
| EC number | 2.1.1.8 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Wikidata | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||


Histamine N-methyltransferase (HNMT) is a cytoplasmic protein encoded by the HNMT gene in humans. It belongs to the methyltransferases superfamily of enzymes and plays a crucial role in the inactivation of histamine, a biogenic amine involved in various physiological processes. Methyltransferases are present in every life form including archaeans, with 230 families of methyltransferases found across species.
Specifically, HNMT transfers a methyl (CH3) group from S-adenosyl-L-methionine (SAM-e) to histamine, forming an inactive metabolite called Nτ-methylhistamine, in a chemical reaction called Nτ-methylation. In mammals, HNMT operates alongside diamine oxidase (DAO) as the only two enzymes responsible for histamine metabolism; however, what sets HNMT apart is its unique presence within the central nervous system (CNS), where it governs intracellular histaminergic neurotransmission. By degrading and regulating levels of intracellular histamine specifically within the CNS, HNMT ensures the proper functioning of neural pathways related to arousal, sleep-wake cycles, and other essential brain functions.
Research on knockout mice lacking the Hnmt gene has revealed that the absence of this enzyme leads to increased brain histamine concentrations and behavioral changes such as heightened aggression and disrupted sleep patterns. These findings highlight the critical role played by HNMT in maintaining normal brain function through precise regulation of neuronal signaling involving histamine. Genetic variants affecting HNMT activity have also been implicated in various neurological disorders like Parkinson's disease and attention deficit disorder, still, as of 2023, it remains unclear whether alterations in HNMT are primary causes or secondary effects of these conditions.
Gene
Histamine N-methyltransferase is encoded by a single gene, called HNMT, which has been mapped to chromosome 2 in humans.[5]
Three transcript variants have been identified for this gene in humans, which produce different protein isoforms[6][5] due to alternative splicing, which allows a single gene to code for multiple proteins by including or excluding particular exons of a gene in the final mRNA produced from that gene.[7][8] Of those isoforms, only one has histamine-methylating activity.[6]
In the human genome, six exons from the 50-kb HNMT contribute to forming a unique mRNA species, approximately 1.6 kb in size. This mRNA is then translated into the cytosolic enzyme histamine N-methyltransferase, comprising 292 amino acids, of which 130 amino acids are a conserved sequence.[9][10] HNMT does not have promoter cis-elements, such as TATA and CAAT boxes.[11] As of 2023, this human gene is still insufficiently characterized.[11]
Protein
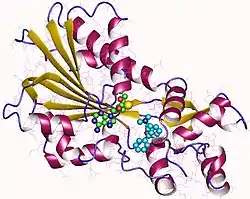
The crystallographic structure (three-dimensional structure) of human HNMT protein was first described in 2001 as a 33kDa monomeric protein consisting of two structural domains.[12][13][11]
The first domain, called the "MTase domain", contains the active site where methylation occurs. It has a classic fold found in many other methyltransferases, and consists of a seven-stranded beta-sheet surrounded by three helices on each side. This domain binds to its cofactor, S-adenosyl-L-methionine (SAM-e), which provides the methyl group for Nτ-methylation reactions.[12][13]
The second domain, called the "substrate binding domain", interacts with histamine, contributing to its binding to the enzyme molecule. This domain is connected to the MTase domain and forms a separate region. It includes an antiparallel beta sheet along with additional alpha helices and 310 helices.[12][13]
Species
Histamine N-methyltransferase belongs to methyltransferases, a superfamily of enzymes present in every life form,[10] including archaeans.[14]
These enzymes catalyze methylation, which is a chemical process that involves the addition of a methyl group to a molecule, which can affect its biological function.[10][13][15]
To facilitate methylation, methyltransferases transfer a methyl group (-CH3) from a cosubstrate (donor) to a substrate molecule (acceptor), leading to the formation of a methylated molecule.[10][13]
Most methyltransferases use S-adenosyl-L-methionine (SAM-e) as a donor, converting it into S-adenosyl-L-homocysteine (SAH).[10][13]
In various species, members of the methyltransferase superfamily of enzymes methylate a wide range of molecules, including small molecules, proteins, nucleic acids, and lipids. These enzymes are involved in numerous cellular processes such as signaling, protein repair, chromatin regulation, and gene regulation. More than 230 families of methyltransferases have been described in various species.[10][16]
This specific protein, histamine N-methyltransferase, is found in vertebrates, including mammals, birds, reptiles, amphibians, and fishes, but not in invertebrates and plants.[17][9][18]
The cDNA of Hnmt was initially cloned from a rat kidney and has since been cloned from human, mouse, and guinea pig sources.[9]
Human HNMT shares 55.37% similarity with that of zebrafish, 86.76% with that of mouse, 90.53% with that of dog, and 99.54% with that of chimpanzee.[18][19] Moreover, expressed sequence tags from cow, pig, and gorilla, as well as genome survey sequences from pufferfish, also exhibit strong similarity to human HNMT, suggesting that it is a highly conserved protein among vertebrates.[12]
To understand the role of histamine N-methyltransferase in brain function, researchers have studied Hnmt-deficient (knockout) mice.[20][17][21] Scientists discovered that disrupting the gene led to a significant rise in histamine levels in the mouse brain that highlighted the role of the gene in the brain's histamine system and suggested that HNMT genetic variations in humans could be linked to brain disorders.[17]
Tissue and subcellular distribution
On subcellular distribution, histamine N-methyltransferase protein in humans is mainly localized to the nucleoplasm (which is an organelle, i.e., a subunit of a cell) and cytosol (which is the intracellular fluid, i.e., a fluid inside cells). In addition, it is localized to the centrosome (another organelle).[22]
In humans, the protein is present in many tissues and is most abundantly expressed in the brain, thyroid gland, bronchus, duodenum, liver, gallbladder, kidney, and skin.[23][17]
Function

The function of the HNMT enzyme is histamine metabolism by ways of Nτ-methylation using S-adenosyl-L-methionine (SAM-e) as the methyl donor, producing Nτ-methylhistamine, which, unless excreted, can be further processed by monoamine oxidase B (MAOB) or by DAO. Methylated histamine metabolites are excreted with urine.[12][13][11]
In mammals, histamine is metabolized by two major pathways: oxidative deamination via DAO, encoded by the AOC1 gene, and Nτ-methylation via histamine N-methyltransferase.[11]
HNMT and DAO are two enzymes that play distinct roles in histamine metabolism. DAO is primarily responsible for metabolizing histamine in extracellular (outside cells) fluids, which include interstitial fluid (fluid surrounding cells) and blood plasma.[17][26] Such histamine can be exogenous (from food or intestinal flora) or endogenous (released from granules of mast cells and basophils, such as during allergic reactions).[27] DAO is predominantly expressed in the cells of the intestinal epithelium and placenta, but not in the central nervous system (CNS).[28][29][17] In contrast, HNMT is expressed in CNS and involved in the metabolism of intracellular (inside cells) histamine, which is primarily endogenous and persistently present. HNMT operates in the cytosol, which is the fluid inside cells.[17] Histamine is required to be carried into the cytosol through transporters[30] such as plasma membrane monoamine transporter (SLC29A4) or organic cation transporter 3 (SLC22A3).[17] HNMT enzyme is found in cells of diverse tissues: neurons and glia, brain, kidneys, liver, bronchi, large intestine, ovary, prostate, spinal cord, spleen, and trachea, etc.[25][31][29][17] While DAO is primarily found in the intestinal epithelium, HNMT is present in a wider range of tissues throughout the body. This difference in location also requires different transport mechanisms for histamine to reach each enzyme, reflecting the distinct roles of these enzymes in histamine metabolism.[17] Another distinction between HNMT and DAO lies in their substrate specificity. While HNMT has a strong preference for histamine, DAO can metabolize other biogenic amines such as putrescine and cadaverine, albeit with a preference for histamine. Both DAO and HNMT exhibit comparable affinities towards histamine.[32]
In the brain of mammals, histamine neurotransmitter activity is controlled by HNMT, since DAO is not present in the CNS.[5] Consequently, the deactivation of histamine via HNMT represents the sole mechanism for ending neurotransmission within the mammalian CNS.[25] This highlights the key role of HNMT for the histamine system of the brain and the brain function in general.[17]
Clinical significance
Role in health
Histamine has important roles in human physiology as both a hormone and a neurotransmitter. As a hormone, it is involved in the inflammatory response and itching. It regulates physiological functions in the gut and acts on the brain, spinal cord, and uterus. As a neurotransmitter, histamine promotes arousal and regulates the sleep-wake cycle. It also affects vasodilation, fluid production in tissues like the nose and eyes, gastric acid secretion, sexual function, and immune responses.[33][34]
HNMT plays a crucial role in maintaining the proper balance of histamine in the human body. HNMT is responsible for the breakdown and metabolism of histamine, converting it into an inactive metabolite, Nτ-methylhistamine,[33][34] which inhibits HNMT expression in a negative feedback loop.[35] By metabolizing histamine, HNMT helps prevent excessive levels of histamine from accumulating in various tissues and organs. This enzymatic activity ensures that histamine remains at appropriate levels to carry out its physiological functions without causing unwanted effects or triggering allergic reactions. In the central nervous system, HNMT plays an essential role in degrading histamine, where it acts as a neurotransmitter, since HNMT is the only enzyme in the body that can metabolize histamine in CNS, ending its neurotransmitter activity.[33][34]
HNMT also plays a role in the airway response to harmful particles,[36] which is the body's physiological reaction to immune allergens, bacteria, or viruses in the respiratory system. Histamine is stored in granules in mast cells, basophils, and in the synaptic vesicles of histaminergic neurons of the airways. When exposed to immune allergens or harmful particles, histamine is released from these storage granules and quickly diffuses into the surrounding tissues. However, the released histamine needs to be rapidly deactivated for proper regulation, which is a function of HNMT.[37][38]
Histamine intolerance
Histamine intolerance (HIT) is a disorder characterized by impaired histamine metabolism, resulting in the excess of active histamine in the body. HIT is often misdiagnosed due to its nonspecific clinical manifestations, which can include itching, flushing, edema, gastrointestinal symptoms such as abdominal pain and diarrhea, respiratory symptoms like runny nose and nasal congestion, and other symptoms like dizziness and headache.[39]
In the case of flawed HNMT activity, the most affected organs are the brain, liver, and mucous membrane of the bronchus. Consequently, flawed HNMT activity can lead to chronic forms of HIT. For instance, the main symptoms of histamine intolerance (HIT) within the nervous system, which are associated with flawed HNMT activity, are anxiety, dizziness, fatigue, insomnia, myoclonic twitching, and unrest. Overall, the symptoms of flawed HNMT activity are typical of symptoms of HIT, including allergic rhinitis, urticaria (hives), and peptic ulcer disease.[17]
The exact mechanisms by which deficiency or impaired activity of HNMT causes HIT are not fully understood but may involve genetic factors. Variations in genes encoding for HNMT have been identified as potential contributors to increased susceptibility to allergic diseases and more severe courses of conditions like atopic dermatitis and allergic rhinitis. While deficiencies in DAO are considered the primary cause of HIT, variations in both DAO and HNMT genes could play a role in its development. The interplay between these enzymes influences how effectively histamine is broken down and cleared from the body.[39][40]
Activity measurements
The activity of HNMT, unlike that of DAO, cannot be measured by blood (serum) analysis.[41][42][43]
Organs that produce DAO continuously release it into the bloodstream. DAO is stored in vesicular structures associated with the plasma membrane in epithelial cells.[29] As a result, serum DAO activity can be measured for diagnosing histamine intolerance, but not HNMT. This is because HNMT is primarily found within the cells of internal organs like the brain or liver and is not released to the bloodstream. Measuring intracellular HNMT directly is challenging. Therefore, diagnosis of HNMT activity is typically done indirectly by testing for known genetic variants.[29]
Genetic variants
There is a genetic variant, registered in The Single Nucleotide Polymorphism Database (dbSNP) as rs11558538, found in 10% of the population worldwide,[44] which means that the T allele presents at position 314 of HNMT instead of a usual C allele (c.314C>T). This variant causes the protein to be synthesized with threonine (Thr) replaced with isoleucine (Ile) at position 105 (p.Thr105Ile, T105I). This variant is described as loss-of-function allele reducing HNMT activity, and is associated with diseases, typical for histamine intolerance, such as asthma, allergic rhinitis, and atopic eczema (atopic dermatitis). For individuals with this variant, the intake of HNMT inhibitors, which hamper enzyme activity, and histamine liberators, which release histamine from the granules of mast cells and basophils, could potentially influence their histamine levels.[45][46] Still, this genetic variant is associated with a reduced risk of Parkinson's disease.[47][48][13]
Experiments involving HNMT knockout mice have shown that a deficiency in HNMT indeed leads to increased brain histamine concentrations, resulting in heightened aggressive behaviors and disrupted sleep-wake cycles in these mice. In humans, genetic variants that affect HNMT activity have been implicated in various brain disorders, such as Parkinson's disease and attention deficit disorder, but it remains unclear whether these alterations in HNMT are a primary cause or secondary effect of these conditions. Additionally, reduced histamine levels in cerebrospinal fluid have been consistently reported in patients with narcolepsy and other conditions characterized by excessive daytime sleepiness. The association between HNMT polymorphisms and gastrointestinal diseases is still uncertain. While mild polymorphisms can lead to diseases such as asthma and inflammatory bowel disease, they may also reduce the risk of brain disorders like Parkinson's disease. On the other hand, severe mutations in HNMT can result in intellectual disability. Despite these findings, the role of HNMT in human health is not fully understood and continues to be an active area of research.[17][25]
Inhibitors
The following substances are known to be HNMT inhibitors: amodiaquine, chloroquine, dimaprit, etoprine, metoprine, quinacrine, SKF-91488, tacrine, and diphenhydramine.[49][50][51] HNMT inhibitors may increase histamine levels in peripheral tissues and cause of aggravate conditions associated with histamine excess, such as allergic rhinitis, urticaria, and peptic ulcer disease.[17] As of 2023, the effect of HNMT inhibitors on brain function is not yet fully understood. Research suggests that using new inhibitors of HNMT to increase the levels of histamine in the brain could potentially contribute to improvements in the treatment of brain disorders.[49][50][51]
Methamphetamine overdose
HNMT could be a potential target for the treatment of symptoms of methamphetamine overdose.[52] It is a central nervous system stimulant, which can be abused up to the lethal consequences: numerous deaths related to methamphetamine overdoses have been reported.[53][54] The reasoning behind this is that such overdose often leads to behavioral abnormalities, and it has been observed that elevated levels of histamine in the brain can attenuate these methamphetamine-induced behaviors. Therefore, by targeting HNMT, it might be possible to increase the levels of histamine in the brain, which could, in turn, help to mitigate the effects of a METH overdose. This effect could be achieved by using HNMT inhibitors. Studies predict that one such inhibitor can be metoprine, which crosses the blood-brain barrier and can potentially increase brain histamine levels by inhibiting HNMT, still, As of 2023, treatment of methamphetamine overdose by HNMT inhibitors is still an area of research.[52]
Nτ-methylhistamine
Nτ-methylhistamine (NMH), also known as 1-methylhistamine, is a product of Nτ-methylation of histamine in a reaction catalyzed by the HNMT enzyme.[24][12][13]
NMH is considered a biologically inactive metabolite of histamine.[55][56][57] NMH is excreted in the urine and can be measured to estimate the amounts of active histamine in the body.[58] While NMH has some biological activity on its own, it is much weaker than histamine. It can bind to histamine receptors, still, NMH has a lower affinity and efficacy than histamine for these receptors, meaning that it binds less strongly and activates them less effectively. Depending on the receptor subtype and the tissue context, NMH may act as a partial agonist or an antagonist for some histamine receptors. NMH may have some modulatory effects on histamine signaling, but it is unlikely to cause significant allergic or inflammatory reactions by itself. NMH may also serve as a feedback mechanism to regulate histamine levels and prevent excessive histamine release.[59] Still, NMT, being a produce in a reaction catalyzed by HNMT, may inhibit expression of HNMT in a negative feedback loop.[35]
Urinary NMH can be measured in clinical settings when systemic mastocytosis is suspected. Systemic mastocytosis and anaphylaxis are typically associated with at least a two-fold increase in urinary NMH levels, which are also increased in patients taking monoamine oxidase inhibitors and in patients on histamine-rich diets.[58]
References
- 1 2 3 GRCh38: Ensembl release 89: ENSG00000150540 - Ensembl, May 2017
- 1 2 3 GRCm38: Ensembl release 89: ENSMUSG00000026986 - Ensembl, May 2017
- ↑ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ↑ "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- 1 2 3
 This article incorporates public domain material from "HNMT Histamine N-methyltransferase". Reference Sequence collection. National Center for Biotechnology Information. Retrieved 30 November 2020.
This article incorporates public domain material from "HNMT Histamine N-methyltransferase". Reference Sequence collection. National Center for Biotechnology Information. Retrieved 30 November 2020. In mammals, histamine is metabolized by two major pathways: N(tau)-methylation via histamine N-methyltransferase and oxidative deamination via diamine oxidase. This gene encodes the first enzyme which is found in the cytosol and uses S-adenosyl-L-methionine as the methyl donor. In the mammalian brain, the neurotransmitter activity of histamine is controlled by N(tau)-methylation as diamine oxidase is not found in the central nervous system. A common genetic polymorphism affects the activity levels of this gene product in red blood cells. Multiple alternatively spliced transcript variants that encode different proteins have been found for this gene.
This article incorporates text from this source, which is in the public domain. - 1 2 "UniProt HNMT isoforms".
- ↑ Marasco LE, Kornblihtt AR (April 2023). "The physiology of alternative splicing". Nat Rev Mol Cell Biol. 24 (4): 242–254. doi:10.1038/s41580-022-00545-z. PMID 36229538. S2CID 252896843.
- ↑ Rogalska ME, Vivori C, Valcárcel J (April 2023). "Regulation of pre-mRNA splicing: roles in physiology and disease, and therapeutic prospects". Nat Rev Genet. 24 (4): 251–269. doi:10.1038/s41576-022-00556-8. PMID 36526860. S2CID 254809593.
- 1 2 3 Barnes WG, Grinde E, Crawford DR, Herrick-Davis K, Hough LB (January 2004). "Characterization of a new mRNA species from the human histamine N-methyltransferase gene". Genomics. 83 (1): 168–71. doi:10.1016/s0888-7543(03)00236-2. PMID 14667820.
- 1 2 3 4 5 6 "InterPro". www.ebi.ac.uk. Retrieved 28 November 2023.
- 1 2 3 4 5 Moya-García AA, Pino-Ángeles A, Sánchez-Jiménez F, Urdiales JL, Medina MÁ (March 2021). "Histamine, Metabolic Remodelling and Angiogenesis: A Systems Level Approach". Biomolecules. 11 (3): 415. doi:10.3390/biom11030415. PMC 8000605. PMID 33799732.
- 1 2 3 4 5 6 Horton JR, Sawada K, Nishibori M, Zhang X, Cheng X (September 2001). "Two polymorphic forms of human histamine methyltransferase: structural, thermal, and kinetic comparisons". Structure. 9 (9): 837–49. doi:10.1016/s0969-2126(01)00643-8. PMC 4030376. PMID 11566133.
- 1 2 3 4 5 6 7 8 9 10 Li J, Sun C, Cai W, Li J, Rosen BP, Chen J (2021). "Insights into S-adenosyl-l-methionine (SAM)-dependent methyltransferase related diseases and genetic polymorphisms". Mutat Res Rev Mutat Res. 788: 108396. doi:10.1016/j.mrrev.2021.108396. PMC 8847900. PMID 34893161.
- ↑ Lee YH, Ren D, Jeon B, Liu HW (September 2023). "S-Adenosylmethionine: more than just a methyl donor". Nat Prod Rep. 40 (9): 1521–1549. doi:10.1039/d2np00086e. PMC 10491745. PMID 36891755.
- 1 2 Chen NH, Djoko KY, Veyrier FJ, McEwan AG (2016). "Formaldehyde Stress Responses in Bacterial Pathogens". Front Microbiol. 7: 257. doi:10.3389/fmicb.2016.00257. PMC 4776306. PMID 26973631.
- ↑ Schubert HL, Blumenthal RM, Cheng X (1 June 2003). "Many paths to methyltransfer: a chronicle of convergence". Trends in Biochemical Sciences. 28 (6): 329–335. doi:10.1016/S0968-0004(03)00090-2. ISSN 0968-0004. PMC 2758044. PMID 12826405.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 Yoshikawa T, Nakamura T, Yanai K (February 2019). "N-Methyltransferase in the Brain". International Journal of Molecular Sciences. 20 (3): 737. doi:10.3390/ijms20030737. PMC 6386932. PMID 30744146.
- 1 2 "HNMT Gene - GeneCards | HNMT Protein | HNMT Antibody".
- ↑ This article incorporates public domain material from "HNMT histamine N-methyltransferase [Danio rerio (Zebrafish)] - Gene - NCBI". Reference Sequence collection. National Center for Biotechnology Information.
- ↑ Naganuma F, Nakamura T, Yoshikawa T, Iida T, Miura Y, Kárpáti A, Matsuzawa T, Yanai A, Mogi A, Mochizuki T, Okamura N, Yanai K (November 2017). "Histamine N-methyltransferase regulates aggression and the sleep-wake cycle". Sci Rep. 7 (1): 15899. Bibcode:2017NatSR...715899N. doi:10.1038/s41598-017-16019-8. PMC 5698467. PMID 29162912.
- ↑ Ogasawara M, Yamauchi K, Satoh Y, Yamaji R, Inui K, Jonker JW, Schinkel AH, Maeyama K (May 2006). "Recent advances in molecular pharmacology of the histamine systems: organic cation transporters as a histamine transporter and histamine metabolism". J Pharmacol Sci. 101 (1): 24–30. doi:10.1254/jphs.fmj06001x6. PMID 16648665.
- ↑ "Subcellular - HNMT - the Human Protein Atlas".
- ↑ "Tissue expression of HNMT - Summary - the Human Protein Atlas".
- 1 2 "Reaction for Histamine N-methyltransferase [EC:2.1.1.8]". KEGG: Kyoto Encyclopedia of Genes and Genomes. R02155.
- 1 2 3 4 Verhoeven WM, Egger JI, Janssen PK, van Haeringen A (December 2020). "Adult male patient with severe intellectual disability caused by a homozygous mutation in the HNMT gene". BMJ Case Rep. 13 (12): e235972. doi:10.1136/bcr-2020-235972. PMC 7735107. PMID 33310825.
- ↑ "Extracellular fluid | Definition, Examples, Function, & Facts | Britannica".
- ↑ Hakl R, Litzman J (2023). "Histamine intolerance". Vnitr Lek. 69 (1): 37–40. doi:10.36290/vnl.2023.005. PMID 36931880. S2CID 257604532.
- ↑ Maintz L, Schwarzer V, Bieber T, van der Ven K, Novak N (2008). "Effects of histamine and diamine oxidase activities on pregnancy: a critical review". Hum Reprod Update. 14 (5): 485–95. doi:10.1093/humupd/dmn014. PMID 18499706.
- 1 2 3 4 Maintz L, Novak N (May 2007). "Histamine and histamine intolerance". The American Journal of Clinical Nutrition. 85 (5): 1185–1196. doi:10.1093/ajcn/85.5.1185. PMID 17490952.
- ↑ Yoshikawa T, Yanai K (2017). "Histamine Clearance Through Polyspecific Transporters in the Brain". Handb Exp Pharmacol. Handbook of Experimental Pharmacology. 241: 173–187. doi:10.1007/164_2016_13. ISBN 978-3-319-58192-7. PMID 27679412.
- ↑ Borriello F, Iannone R, Marone G (2017). "Histamine Release from Mast Cells and Basophils". Histamine and Histamine Receptors in Health and Disease. Handbook of Experimental Pharmacology. Vol. 241. Springer. pp. 121–139. doi:10.1007/164_2017_18. ISBN 978-3-319-58192-7. PMID 28332048.
- ↑ Comas-Basté O, Sánchez-Pérez S, Veciana-Nogués MT, Latorre-Moratalla M, Vidal-Carou MDC (August 2020). "Histamine Intolerance: The Current State of the Art". Biomolecules. 10 (8): 1181. doi:10.3390/biom10081181. PMC 7463562. PMID 32824107.
- 1 2 3 Lieberman P (February 2011). "The basics of histamine biology". Ann Allergy Asthma Immunol. 106 (2 Suppl): S2–5. doi:10.1016/j.anai.2010.08.005. PMID 21277530.
- 1 2 3 Mochizuki T (2022). "Histamine as an Alert Signal in the Brain". Curr Top Behav Neurosci. Current Topics in Behavioral Neurosciences. 59: 413–425. doi:10.1007/7854_2021_249. ISBN 978-3-031-16996-0. PMID 34448132. S2CID 237329317.
- 1 2 Peters LJ, Kovacic JP (August 2009). "Histamine: metabolism, physiology, and pathophysiology with applications in veterinary medicine". J Vet Emerg Crit Care (San Antonio). 19 (4): 311–28. doi:10.1111/j.1476-4431.2009.00434.x. PMID 25164630.
- ↑ "UniProt HNMT".
- ↑ Schwartz JC, Arrang JM, Garbarg M, Pollard H, Ruat M (January 1991). "Histaminergic transmission in the mammalian brain". Physiol Rev. 71 (1): 1–51. doi:10.1152/physrev.1991.71.1.1. PMID 1846044.
- ↑ Sande CJ, Njunge JM, Mwongeli Ngoi J, Mutunga MN, Chege T, Gicheru ET, Gardiner EM, Gwela A, Green CA, Drysdale SB, Berkley JA, Nokes DJ, Pollard AJ (May 2019). "Airway response to respiratory syncytial virus has incidental antibacterial effects". Nat Commun. 10 (1): 2218. Bibcode:2019NatCo..10.2218S. doi:10.1038/s41467-019-10222-z. PMC 6525170. PMID 31101811.
- 1 2 Zhao Y, Zhang X, Jin H, Chen L, Ji J, Zhang Z (March 2022). "Histamine Intolerance-A Kind of Pseudoallergic Reaction". Biomolecules. 12 (3): 454. doi:10.3390/biom12030454. PMC 8945898. PMID 35327646.
- ↑ Hrubisko M, Danis R, Huorka M, Wawruch M (June 2021). "Histamine Intolerance-The More We Know the Less We Know. A Review". Nutrients. 13 (7): 2228. doi:10.3390/nu13072228. PMC 8308327. PMID 34209583.
- ↑ Heidari A, Tongsook C, Najafipour R, Musante L, Vasli N, Garshasbi M, Hu H, Mittal K, McNaughton AJ, Sritharan K, Hudson M, Stehr H, Talebi S, Moradi M, Darvish H, Arshad Rafiq M, Mozhdehipanah H, Rashidinejad A, Samiei S, Ghadami M, Windpassinger C, Gillessen-Kaesbach G, Tzschach A, Ahmed I, Mikhailov A, Stavropoulos DJ, Carter MT, Keshavarz S, Ayub M, Najmabadi H, Liu X, Ropers HH, Macheroux P, Vincent JB (October 2015). "Mutations in the histamine N-methyltransferase gene, HNMT, are associated with nonsyndromic autosomal recessive intellectual disability". Hum Mol Genet. 24 (20): 5697–710. doi:10.1093/hmg/ddv286. PMC 4581600. PMID 26206890.
- ↑ Scott MC, Guerciolini R, Szumlanski C, Weinshilboum RM (March 1991). "Mouse kidney histamine N-methyltransferase: assay conditions, biochemical properties and strain variation". Agents Actions. 32 (3–4): 194–202. doi:10.1007/BF01980873. PMID 1907425. S2CID 35519684.
- ↑ McClain JL, Morales-Soto W, Gonzales J, Parmar V, Demireva EY, Gulbransen BD (November 2023). "Sexually Dimorphic Effects of Histamine Degradation by Enteric Glial Histamine N-Methyltransferase (HNMT) on Visceral Hypersensitivity". Biomolecules. 13 (11): 1651. doi:10.3390/biom13111651. PMC 10669271. PMID 38002333.
- ↑ This article incorporates public domain material from "rs11558538 RefSNP Report - dbSNP - NCBI". Reference Sequence collection. National Center for Biotechnology Information.
- ↑ García-Martín E, Ayuso P, Martínez C, Blanca M, Agúndez JA (May 2009). "Histamine pharmacogenomics". Pharmacogenomics. 10 (5): 867–83. doi:10.2217/pgs.09.26. PMID 19450133.
- ↑ Dvornikova KA, Platonova ON, Bystrova EY (June 2023). "Inflammatory Bowel Disease: Crosstalk between Histamine, Immunity, and Disease". Int J Mol Sci. 24 (12): 9937. doi:10.3390/ijms24129937. PMC 10297875. PMID 37373085.
- ↑ Lu Y, Dong CZ, Bao D, Zhong C, Liu K, Chen L, Wang W, Yang B (November 2022). "The Thr105Ile Variant (rs11558538) of the Histamine N-methyltransferase Gene may be associated with Reduced Risk of Parkinson Disease: A Meta-analysis". Genet Test Mol Biomarkers. 26 (11): 543–549. doi:10.1089/gtmb.2021.0299. PMID 36378841. S2CID 253551556.
- ↑ Jiménez-Jiménez FJ, Alonso-Navarro H, García-Martín E, Agúndez JA (July 2016). "Thr105Ile (rs11558538) polymorphism in the histamine N-methyltransferase (HNMT) gene and risk for Parkinson disease: A PRISMA-compliant systematic review and meta-analysis". Medicine (Baltimore). 95 (27): e4147. doi:10.1097/MD.0000000000004147. PMC 5058861. PMID 27399132.
- 1 2 Flores-Clemente C, Nicolás-Vázquez MI, Mera Jiménez E, Hernández-Rodríguez M (September 2021). "Inhibition of Astrocytic Histamine N-Methyltransferase as a Possible Target for the Treatment of Alzheimer's Disease". Biomolecules. 11 (10): 1408. doi:10.3390/biom11101408. PMC 8533269. PMID 34680041.
- 1 2 Horton JR, Sawada K, Nishibori M, Cheng X (October 2005). "Structural basis for inhibition of histamine N-methyltransferase by diverse drugs". J Mol Biol. 353 (2): 334–344. doi:10.1016/j.jmb.2005.08.040. PMC 4021489. PMID 16168438.
- 1 2 Pavadai L (2012). "Pharmacophore modeling, virtual screening and docking studies to identify novel HNMT inhibitors". Journal of the Taiwan Institute of Chemical Engineers. 43 (4): 493–503. doi:10.1016/j.jtice.2012.01.004.
- 1 2 Kitanaka J, Kitanaka N, Hall FS, Uhl GR, Takemura M (2016). "Brain Histamine N-Methyltransferase As a Possible Target of Treatment for Methamphetamine Overdose". Drug Target Insights. 10: 1–7. doi:10.4137/DTI.S38342. PMC 4777238. PMID 26966348.
- ↑ "Meth Overdose Symptoms, Effects & Treatment | BlueCrest". Bluecrest Recovery Center. 17 June 2019. Archived from the original on 16 January 2021. Retrieved 8 October 2020.
- ↑ "Overdose Death Rates". National Institute on Drug Abuse. 29 January 2021. Archived from the original on 25 January 2018. Retrieved 8 October 2020.
- ↑ Maslinski S, Schippert B, Kovar KA, Sewing KF (1977). "Methylation of histamine in the gastric mucosa". Digestion. 15 (6): 497–505. doi:10.1159/000198040. PMID 913915.
- ↑ Murray S, Taylor GW, Karim QN, Bliss P, Calam J (November 2000). "N alpha-methylhistamine: association with Helicobacter pylori infection in humans and effects on gastric acid secretion". Clin Chim Acta. 301 (1–2): 181–92. doi:10.1016/s0009-8981(00)00357-0. PMID 11020472.
- ↑ Grassmann S, Apelt J, Ligneau X, Pertz HH, Arrang JM, Ganellin CR, Schwartz JC, Schunack W, Stark H (October 2004). "Search for histamine H(3) receptor ligands with combined inhibitory potency at histamine N-methyltransferase: omega-piperidinoalkanamine derivatives". Arch Pharm (Weinheim). 337 (10): 533–45. doi:10.1002/ardp.200400897. PMID 15476285. S2CID 19755327.
- 1 2 Lewiecki M (2013). "Evaluation of the Patient at Risk for Osteoporosis". Chapter 63 - Evaluation of the Patient at Risk for Osteoporosis. Academic Press. pp. 1481–1504. doi:10.1016/B978-0-12-415853-5.00063-7. ISBN 978-0-12-415853-5.
- ↑ Mohammed T (2010). "Biological and Pharmacological Aspects of Histamine Receptors and Their Ligands". Biomedical Aspects of Histamine. Springer. pp. 61–100. doi:10.1007/978-90-481-9349-3_4. ISBN 978-90-481-9348-6.
This article incorporates text from the United States National Library of Medicine, which is in the public domain.
External links
- Histamine+N-Methyltransferase at the U.S. National Library of Medicine Medical Subject Headings (MeSH)
- PDBe-KB provides an overview of all the structure information available in the PDB for human histamine N-methyltransferase