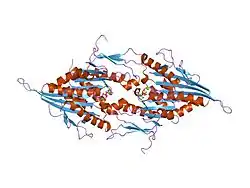


Kinesin-like protein KIF11 is a molecular motor protein that is essential in mitosis. In humans it is coded for by the gene KIF11.[5][6] Kinesin-like protein KIF11 is a member of the kinesin superfamily, which are nanomotors that move along microtubule tracks in the cell. Named from studies in the early days of discovery, it is also known as Kinesin-5,[7] or as BimC, Eg5 or N-2, based on the founding members of this kinesin family.
Currently, there are over 70 different eukaryotic kinesin-5 proteins identified by sequence similarity. Members of this protein family are known to be involved in various kinds of spindle dynamics and essential for mitosis. The function of this gene product includes chromosome positioning, centrosome separation and establishing a bipolar spindle during cell mitosis.[7] The human kinesin-5 protein has been actively studied for its role in mitosis and its potential as a therapeutic target for cancer.
Function
KIF11 (also known as kinesin-5 and Eg5) is a homotetramer which cross-links anti-parallel microtubules in the mitotic spindle to maintain spindle bipolarity.[8][9][10][11] The motor domain or motor head is at the N-terminus and performs ATP hydrolysis and binds to microtubules. Kinesin-5 motors assemble into a bipolar homotetrameric structure that is capable of sliding apart bundles of anti-parallel oriented microtubules.[9][12][13] This motor is essential for mitosis in most organisms, wherein it participates in the self-assembly of the microtubule-based mitotic spindle, but is not otherwise required for cell viability. The motor may also play a role in the proper development of mammalian neuronal processes, including growth cone navigation and elongation.[14][15]
Function in mitosis
In most eukaryotic cells, Kinesin-5 is thought to form cross-bridges between pairs of oppositely oriented microtubules in prophase and prometaphase and drives apart duplicated centrosomes during the formation of the mitotic spindle.[9][13][16] This permits the establishment of a steady-state bipolar microtubule spindle structure.
Loss of Kinesin-5 function from the onset of mitosis in most eukaryotic organisms examined, including animals, plants, and fungi, results in catastrophic failure of mitosis.[17][18][19][20][21][22] This motor's function is crucial during the onset of mitosis, wherein its loss of function results in the collapse, or inversion, of the spindle poles leaving centrally positioned centrosome pairs flanked by a radial array of microtubules with peripheral condensed chromosomes. The one exception to this effect is mitosis within the nematode, C. elegans, in which Kinesin-5 is not strictly essential for mitosis, but nonetheless has considerable impact on the overall fidelity of cell division.[23]
The discovery of small chemical inhibitors of human Kinesin-5 through a pioneering in vitro phenotypic screening on cancer cell lines has led to both the development of new anticancer therapeutic agents, and to novel tools to probe the mechanism of microtubule motor proteins.[22][24] This toolkit of allosteric inhibitors has been used to probe the specific role of Kinesin-5 in mitotic spindle assembly [25] as well as fine dissection of motor domain function.[26][27][28][29][30] Through this work it was found that, in mammalian cells, Kinesin-5 is required for the initial assembly of the mitotic spindle during prophase and prometaphase, but is dispensable to traverse subsequent anaphase during a round of mitosis.[8][25] Also, the binding of the Kinesin-5 inhibitors to an allosteric site on the motor interrupts the mechanism by which this enzyme converts the chemical energy of ATP hydrolysis into the mechanical work of moving microtubules, thus providing insight on how this enzyme works.
There are many models that attempt to explain the self-assembly of the mitotic spindle based upon microtubules as a structural element, and a set of microtubule motors, including Kinesin-5 to move and order them. Many of these models attempt to explain the steady state of the spindle at metaphase based on a predicted balance of motor forces acting in opposition within the spindle microtubules.[31][32] Still, it is not clear whether all the structural elements required for spindle assembly are known, or how the motors, including Kinesin-5, might be regulated in space and time. Such caveats make assessment of such models difficult. Recent data, however, finds that aspects of the 'force balance' model that posit spindle length and stability to be mediated by a balance between the minus-end directed microtubule sliding and plus-end directed microtubule sliding by opposing motors in insect cells, seems not to be the case in mammalian cells.[33] The process of self-assembly of the mitotic spindle remains a major unsolved question in cell biology, and a robust model awaits further details of the regulation and behavior of various microtubule motors and structural elements that compose this machinery.
Function in neurons
Although Kinesin-5 is required in all cells during cell division, it does not appear to play a major role in the metabolism of most non-dividing cells.[21][22] Among non-dividing cells, Kinesin-5 is most enriched within neurons, wherein it decorates the large microtubule bundles extending into axons and dendrites.[22][34] It has been shown, for example, that neurons remain fully viable in the background of a knock-down of Kinesin-5, but that changes in neuronal development and morphogenesis ensue. In developing neurons pharmacological inhibition and siRNA knockdown of KIF11 results in longer axons, more branches, fewer bouts of axon retraction and the inability of growth cones to turn on contact with repulsive substrates.[35][36][37] In migratory neurons, inhibition of KIF11 causes neurons to migrate in a random pattern and form shorter leading processes.[15] KIF11, like KIF15 and KIF23, is thought to act as a restrictor of short microtubules moving bi-directionally along the axon, exerting forces antagonistically to cytoplasmic dynein.[38][39] In mature neurons, KIF11 restricts the movement of short microtubules in dendrites, contributing to the formation of characteristic shape of dendrites.[40] KIF11 is also expressed in adult dorsal root ganglion neurons, although at a much diminished level. In adult neurons It has a similar effect on inhibiting the rate of short microtubule transport so pharmacological inhibition and siRNA knockdown of adult KIF11 may be a potential therapeutic tool for the augmentation of adult axon regeneration.[41] However, a clear in vivo role for Kinesin-5 in neurogenesis remains to be elucidated. Of note is that unusual peripheral neuropathies have not been observed in patients undergoing recent phase I or phase II trials of Kinesin-5 inhibitors for potential anti-cancer therapy.[42][43]
Functional regulation
In 1995, Kinesin-5 was determined to be post-translationally phosphorylated within its C-terminal tail.[8][44] Once Kinesin-5 is phosphorylated at this residue in early prophase, it localizes to the mitotic spindle where it binds to microtubules. An additional phosphosite was identified on the Kinesin-5 tail in 2008, however, only approximately 3% of the total microtubule-associated Kinesin-5 is phosphorylated at this residues.[45] While additional phosphosites or other post-translational modifications within the Kinesin-5 tail, stalk, and motor have been identified,[46][47] no other modifications have been proven as necessary for Kinesin-5 to perform its necessary tasks in mitosis.
Kinesin-5 is also regulated through direct interaction with other proteins. The microtubule-associated protein, TPX2, associates with Kinesin-5 in mitosis. Their interaction is necessary for Kinesin-5 localization to the mitotic spindle, for stabilizing the spindle, and for spindle pole segregation.[48][49] Kinesin-5 has been shown to interact with the dynactin subunit p150Glued[50] as well as many other cell cycle related proteins in vivo and in vitro,[51][52][53] however, additional experimentation is needed to confirm that their association is necessary for Kinesin-5 to function normally.
Molecular mechanism
ATP hydrolysis
Kinesin-5, like all motor proteins, breaks down ATP into ADP and inorganic phosphate, using a water molecule, and converts the chemical energy to force and motion along microtubules. Kinetic experiments reveal rates of how fast intermediate steps in catalysis occur and the most extensive set of studies on Kinesin-5 kinetics has been on the human protein.[54][55] X-ray crystallography, cryo-electron microscopy, and real-time infrared spectroscopy have been used to measure the structure of Kinesin-5 in the different catalytic intermediate states. Changes in the secondary structure, or conformational switching, is required to convert and amplify biochemical changes in the catalytic active site into larger movements necessary for cellular motion.[56][57] For example, the first step of ATP hydrolysis, which is the attack of the terminal phosphate of ATP by a water molecule, had not been observed by x-ray crystallography in any kinesin protein, until recently in Kinesin-5.[58] This crystal structure showed that there was not one, but rather two, water molecules and they are in close association with each other. A two-water catalytic model was proposed and confirmed by an alternate method to track Kinesin-5 catalysis in real-time[59] and in a kinesin protein in a different subfamily.[60] Two-water catalytic models also are proposed in a divergent motor protein, myosin, and observed experimentally in one of its crystal structures.[61][62]
Mechanical Properties
The antiparallel tetrameric organization of the Kinesin-5 family is fundamentally different from the majority of other kinesins that are dimers, such as the well-characterized conventional Kinesin-1 (KIF5B). Conventional kinesin dimerizes in such a manner that the catalytic (head) domains are together on one end of the complex to facilitate hand-over-hand movement along a microtubule that enables long-range, directed transport of cellular cargoes. The unique assembly of Kinesin-5 proteins not only organizes the protein complex for a different cellular function (antiparallel microtubule sliding, described above) but also made it difficult to study the mechanical properties of the motor using the classical experiments that were designed for dimeric kinesins. These obstacles have been overcome by either adapting the original experiments to analyze the tetrameric organization of Kinesin-5, or by working with shorter Kinesin-5 proteins that form dimers like conventional kinesin.
The most striking outcomes of the analysis of Kinesin-5 motility is that it is slow – about 10 times slower than conventional Kinesin-1 – with a velocity in the range of 50 nanometers per second and that it could generate very high levels of mechanical force (7-9 picoNewtons per molecule). These values come from three types of experimental data: microtubule gliding assays, single molecule motility assays, and optical trap assays. In microtubule gliding assays, kinesins are attached to a glass surface and microtubules are laid down over the top. Since the motors are attached to the glass, their motile behavior translates into movement of the microtubule across the anchored kinesins, akin to someone crowd surfing. These experiments gave us the first analysis of Kinesin-5 motility.

By attaching microtubules to the glass surface first, then adding Kinesin-5 with free microtubules in solution, it was possible to adapt the microtubule gliding assays to show that Kinesin-5 can crosslink two microtubules and move them in opposite directions. This experiment showed that Kinesin-5 was indeed capable of carrying out the role that had been proposed for it in mitosis – sliding oppositely oriented microtubules in the mitotic spindle. To study the behavior of individual Kinesin-5 molecules, single molecule motility assays were performed by attaching microtubules to a glass surface, then adding a dilute solution of Kinesin-5 with a fluorophore attached. This experimental setup enables the observer to follow separate Kinesin-5 molecules as they "walk" along the microtubule, providing not only information about velocity, but also about processivity – the ability of a kinesin to take multiple steps along the microtubule without dissociating. Kinesin-5 in this setup has shown bi-directionality. Thus it can "walk" in both direction. The switching of direction is controlled with high precision. In single molecule motility assays, velocities for Kinesin-5 were similar to those seen in microtubule gliding assays, and the motor was observed to be weakly processive.[63][64][65] In optical trap experiments, Kinesin-5 molecules are attached to a bead that can be held in place by a finely focused laser. By moving the bead close to a microtubule, the kinesin can bind to the microtubule and begin stepping, pulling the bead along behind it. Since the bead is being held in place by the trap laser, it acts like a spring and exerts a force that resists the forward movement of the kinesin. This allows for the measurement of the stall force – the maximum amount of force that can be exerted by a motor before it releases from the microtubule. Optical trap experiments showed that Kinesin-5 generates a maximum of 7 picoNewtons of force before releasing, but that its behavior differs from that of other kinesins in that there was no observable plateau phase in which the motor "struggles" at its maximal force generation before letting go.[66][67] Extrapolation of kinetic data suggests that the maximal observed force generated in the optical trap by Kinesin-5 is actually an underestimate and that it theoretically can exert up to 9 picoNewtons of force as a maximum, although further experimental work is required to test this.
Pharmacological inhibitors
Inhibitors of KIF11 have been developed as chemotherapeutic agents in the treatment of cancer. Drugs that specifically inhibit only human Kinesin-5 are alternatives to the taxanes and vinc alkaloids that target microtubules, and thus all cells, and that are currently used clinically. Inhibition of Kinesin-5 causes cells to undergo mitotic arrest, undergo apoptosis and form monoaster spindles.[68] The first KIF11 inhibitor, monastrol was discovered in a chemical screen of a large library of cell permeable compounds.[22][69] Since then, over 100 different chemical classes of allosteric inhibitors have been identified in the scientific literature and they have a wide range in potency against human Kinesin-5.[43][70] Common KIF11 inhibitors include:
The majority of human Kinesin-5 inhibitors are selective, because they bind to a drug 'hot spot', composed of residues from the α2 and α3 helices and a flexible L5 loop on the surface of the motor domain. This L5 loop has high sequence variability amongst the Kinesin-5 orthologs and other kinesins within the superfamily. The L5 loop in human Kinesin-5 closes around the inhibitor and is open in the absence of inhibitor.[74][75] These structural changes are correlated with other changes in the catalytic active site. Other sites of inhibitor binding have been identified in the human Kinesin-5 motor domain.[76][77] For inhibitors that bind to the L5 pocket, the mechanism of inhibition is that they slow ADP release from the catalytic active site[78] and inhibit ATP-dependent directional motion.[79] However, a previously unknown diffusive motion by Kinesin-5 along microtubules was uncovered when monastrol inhibited the motor domain.[80]
Small-molecule inhibitors are not only important tools for understanding nanomotors in cells; they are also have potential for serving as tools in the clinic. Induced by human Kinesin-5 inhibitors, mitotic arrest results in apoptosis in some tumor cell lines[81][82] and human tumor xenograft models.[83] With these promising preclinical studies, ispinesib (SB-715992; Cytokinetics/GSK), SB-743921 from Cytokinetics/GSK,[84] MK-0731 from Merck,[85] filanesib (ARRY-520) (Array BioPharma), and litronesib (LY2523355) (Eli Lilly) have entered into clinical trials.[86][87][88] Although second-generation Kinesin-5 inhibitors have had better success, none have been fully developed and marketed as an anti-cancer treatment.
The role of specific residues in the L5 pocket (L5, α2, and α3) in human Kinesin-5 has been tested,[26][28][89][90] but not yet been systematically explored. The initial goal of these mutation experiments was to determine which residues had greatest pharmacological importance in drug development. For example, mutations in the KIF11 gene convey resistance of mitotic cell lines to inhibitors such as monastrol and STLC.[28][91] For example, point mutations in the inhibitor binding pocket, R119A, D130A, L132A, I136A, L214A and E215A confer resistance to monastrol, while R119A, D130A and L214A mutations confer resistance to STLC. In contrast to the loss-of-function experiments, a gain-of-function experiment using Drosophila Kinesin-5 showed that all L5-directed inhibitors do not allosterically communicate in the same way within the Kinesin-5 motor domain.[30]
A second purpose of mutational studies is to understand how drug resistance in cells is conferred from only changing one residue. These changes in the inhibitor-binding pocket are correlated with structural modification, or twist, of the central beta-sheet of the Kinesin-5 motor domain.[28] In this manner, the L5 loop may be able to directly control nucleotide binding and beta-sheet twist can manipulate the adjacent microtubule-binding site. This may explain how tumor cells rapidly can become drug-resistant to KIF11 inhibitors.
Human mutations
KIF11 mutations have been widely described in cancer, and many trials with KIF11 inhibitors are ongoing.
Clinical significance
Germline mutations in KIF11 cause microcephaly with or without chorioretinopathy, lymphedema, or mental retardation (MCLMR).[92] This syndrome is observed as an autosomal dominant disorder with variable expressivity but can also be sporadic. It is characterized by mild-to-severe microcephaly, often associated with developmental delay, ocular defects and lymphedema, usually on the dorsum of the feet. Phenotypic evaluation of patients (n = 87) revealed microcephaly in 91%, eye anomalies in 72%, intellectual disability in 67% and lymphedema in 47% of the patients. Unaffected carriers were rare (4 out of 87: 5%). Family history is not a requisite for diagnosis; 31% (16 out of 52) were de novo cases. All inherited cases, and 50% of sporadic cases of MCLMR are due to germline KIF11 mutations.[93]
Notes
References
- 1 2 3 GRCh38: Ensembl release 89: ENSG00000138160 - Ensembl, May 2017
- 1 2 3 GRCm38: Ensembl release 89: ENSMUSG00000012443 - Ensembl, May 2017
- ↑ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ↑ "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ↑ "KIF11 - Kinesin-like protein KIF11 - Homo sapiens (Human) - KIF11 gene & protein". www.uniprot.org. Retrieved 10 April 2022.
- ↑ Wojcik EJ, Buckley RS, Richard J, Liu L, Huckaba TM, Kim S (December 2013). "Kinesin-5: Cross-bridging mechanism to targeted clinical therapy". Gene. 531 (2): 133–49. doi:10.1016/j.gene.2013.08.004. PMC 3801170. PMID 23954229.
- 1 2 "Entrez Gene: Kinesin family member 11".
- 1 2 3 Blangy A, Lane HA, d'Hérin P, Harper M, Kress M, Nigg EA (December 1995). "Phosphorylation by p34cdc2 regulates spindle association of human Eg5, a kinesin-related motor essential for bipolar spindle formation in vivo". Cell. 83 (7): 1159–69. doi:10.1016/0092-8674(95)90142-6. PMID 8548803.
- 1 2 3 Kashina AS, Baskin RJ, Cole DG, Wedaman KP, Saxton WM, Scholey JM (January 1996). "A bipolar kinesin". Nature. 379 (6562): 270–2. Bibcode:1996Natur.379..270K. doi:10.1038/379270a0. PMC 3203953. PMID 8538794.
- ↑ Sharp DJ, McDonald KL, Brown HM, Matthies HJ, Walczak C, Vale RD, Mitchison TJ, Scholey JM (January 1999). "The bipolar kinesin, KLP61F, cross-links microtubules within interpolar microtubule bundles of Drosophila embryonic mitotic spindles". J. Cell Biol. 144 (1): 125–38. doi:10.1083/jcb.144.1.125. PMC 2148119. PMID 9885249.
- ↑ Sharp DJ, Yu KR, Sisson JC, Sullivan W, Scholey JM (May 1999). "Antagonistic microtubule-sliding motors position mitotic centrosomes in Drosophila early embryos". Nat. Cell Biol. 1 (1): 51–4. doi:10.1038/9025. PMID 10559864. S2CID 6229447.
- ↑ Cole DG, Saxton WM, Sheehan KB, Scholey JM (1994). "A "slow" homotetrameric kinesin-related motor protein purified from Drosophila embryos". J Biol Chem. 269 (37): 22913–6. doi:10.1016/S0021-9258(17)31593-4. PMC 3201834. PMID 8083185.
- 1 2 Sawin KE, LeGuellec K, Philippe M, Mitchison TJ (October 1992). "Mitotic spindle organization by a plus-end-directed microtubule motor". Nature. 359 (6395): 540–3. Bibcode:1992Natur.359..540S. doi:10.1038/359540a0. PMID 1406972. S2CID 4358461.
- ↑ Ferhat L, Cook C, Chauviere M, Harper M, Kress M, Lyons GE, Baas PW (October 1998). "Expression of the mitotic motor protein Eg5 in postmitotic neurons: implications for neuronal development". J. Neurosci. 18 (19): 7822–35. doi:10.1523/JNEUROSCI.18-19-07822.1998. PMC 6793023. PMID 9742151.
- 1 2 Falnikar A, Tole S, Baas PW (2011). "Kinesin-5, a mitotic microtubule-associated motor protein, modulates neuronal migration". Mol Biol Cell. 22 (9): 1561–74. doi:10.1091/mbc.E10-11-0905. PMC 3084678. PMID 21411631.
- ↑ Acar S, Carlson DB, Budamagunta MS, Yarov-Yarovoy V, Correia JJ, Ninonuevo MR, Jia W, Tao L, Leary JA, Voss JC, Evans JE, Scholey JM (2013). "The bipolar assembly domain of the mitotic motor kinesin-5". Nat Commun. 4 (4): 1343. Bibcode:2013NatCo...4.1343A. doi:10.1038/ncomms2348. PMC 3562449. PMID 23299893.
- ↑ Heck MM, Pereira A, Pesavento P, Yannoni Y, Spradling AC, Goldstein LS (1993). "The kinesin-like protein KLP61F is essential for mitosis in Drosophila". J Cell Biol. 123 (3): 665–79. doi:10.1083/jcb.123.3.665. PMC 2200134. PMID 8227131.
- ↑ Bannigan A, Scheible WR, Lukowitz W, Fagerstrom C, Wadsworth P, Somerville C, Baskin TI (2007). "A conserved role for kinesin-5 in plant mitosis". J Cell Sci. 120 (Pt 16): 2819–27. doi:10.1242/jcs.009506. PMID 17652157.
- ↑ Enos AP, Morris NR (1990). "Mutation of a gene that encodes a kinesin-like protein blocks nuclear division in A. nidulans". Cell. 60 (6): 1019–27. doi:10.1016/0092-8674(90)90350-N. PMID 2138511. S2CID 27420513.
- ↑ Hagan I, Yanagida M (1990). "Novel potential mitotic motor protein encoded by the fission yeast cut7+ gene". Nature. 347 (6293): 563–6. Bibcode:1990Natur.347..563H. doi:10.1038/347563a0. PMID 2145514. S2CID 4234302.
- 1 2 Sawin KE, Mitchison TJ, Wordeman LG (1992). "Evidence for kinesin-related proteins in the mitotic apparatus using peptide antibodies". J Cell Sci. 101 (Pt 2): 303–13. doi:10.1242/jcs.101.2.303. PMID 1629247.
- 1 2 3 4 5 6 Mayer TU, Kapoor TM, Haggarty SJ, King RW, Schreiber SL, Mitchison TJ (1999). "Small molecule inhibitor of mitotic spindle bipolarity identified in a phenotype-based screen". Science. 286 (5441): 971–4. doi:10.1126/science.286.5441.971. PMID 10542155.
- ↑ Bishop JD, Han Z, Schumacher JM (2005). "The Caenorhabditis elegans Aurora B kinase AIR-2 phosphorylates and is required for the localization of a BimC kinesin to meiotic and mitotic spindles". Mol Biol Cell. 16 (2): 742–56. doi:10.1091/mbc.E04-08-0682. PMC 545908. PMID 15548597.
- 1 2 DeBonis S, Skoufias DA, Lebeau L, Lopez R, Robin G, Margolis RL, Wade RH, Kozielski F (2004). "In vitro screening for inhibitors of the human mitotic kinesin Eg5 with antimitotic and antitumor activities". Mol Cancer Ther. 3 (9): 1079–90. doi:10.1158/1535-7163.1079.3.9. PMID 15367702. S2CID 15725001.
- 1 2 3 Kapoor TM, Mayer TU, Coughlin ML, Mitchison TJ (2000). "Probing spindle assembly mechanisms with monastrol, a small molecule inhibitor of the mitotic kinesin". J Cell Biol. 150 (5): 975–88. doi:10.1083/jcb.150.5.975. PMC 2175262. PMID 10973989.
- 1 2 Brier S, Lemaire D, Debonis S, Forest E, Kozielski F (2004). "Identification of the protein binding region of S-trityl-L-cysteine, a new potent inhibitor of the mitotic kinesin Eg5". Biochemistry. 43 (41): 13072–82. doi:10.1021/bi049264e. PMID 15476401.
- ↑ Larson AG, Naber N, Cooke R, Pate E, Rice SE (2010). "The conserved L5 loop establishes the pre-powerstroke conformation of the Kinesin-5 motor, eg5". Biophys J. 98 (11): 2619–27. Bibcode:2010BpJ....98.2619L. doi:10.1016/j.bpj.2010.03.014. PMC 2877332. PMID 20513406.
- 1 2 3 4 Kim ED, Buckley R, Learman S, Richard J, Parke C, Worthylake DK, Wojcik EJ, Walker RA, Kim S (2010). "Allosteric drug discrimination is coupled to mechanochemical changes in the kinesin-5 motor core". J Biol Chem. 285 (24): 18650–61. doi:10.1074/jbc.M109.092072. PMC 2881790. PMID 20299460.
- ↑ Wojcik EJ, Dalrymple NA, Alford SR, Walker RA, Kim S (2004). "Disparity in allosteric interactions of monastrol with Eg5 in the presence of ADP and ATP: a difference FT-IR investigation". Biochemistry. 43 (31): 9939–49. CiteSeerX 10.1.1.495.1844. doi:10.1021/bi048982y. PMID 15287721.
- 1 2 Liu L, Parameswaran S, Liu J, Kim S, Wojcik EJ (2011). "Loop 5-directed compounds inhibit chimeric kinesin-5 motors: implications for conserved allosteric mechanisms". J Biol Chem. 286 (8): 6201–10. doi:10.1074/jbc.M110.154989. PMC 3057856. PMID 21127071.
- ↑ Mogilner A, Craig E (2010). "Towards a quantitative understanding of mitotic spindle assembly and mechanics". J Cell Sci. 123 (Pt 20): 3435–45. doi:10.1242/jcs.062208. PMC 2951465. PMID 20930139.
- ↑ Karsenti E, Vernos I (2001). "The mitotic spindle: a self-made machine". Science. 294 (5542): 543–7. Bibcode:2001Sci...294..543K. doi:10.1126/science.1063488. PMID 11641489. S2CID 32846903.
- ↑ Florian S, Mayer TU (2012). "The functional antagonism between Eg5 and dynein in spindle bipolarization is not compatible with a simple push-pull model". Cell Rep. 1 (5): 408–16. doi:10.1016/j.celrep.2012.03.006. PMID 22832270.
- ↑ Haque SA, Hasaka TP, Brooks AD, Lobanov PV, Baas PW (2004). "Monastrol, a prototype anti-cancer drug that inhibits a mitotic kinesin, induces rapid bursts of axonal outgrowth from cultured postmitotic neurons". Cell Motil Cytoskeleton. 58 (1): 10–6. CiteSeerX 10.1.1.507.5695. doi:10.1002/cm.10176. PMID 14983520.
- ↑ Myers KA, Baas PW (September 2007). "Kinesin-5 regulates the growth of the axon by acting as a brake on its microtubule array". J. Cell Biol. 178 (6): 1081–91. doi:10.1083/jcb.200702074. PMC 2064629. PMID 17846176.
- ↑ Nadar VC, Ketschek A, Myers KA, Gallo G, Baas PW (December 2008). "Kinesin-5 is essential for growth-cone turning". Curr. Biol. 18 (24): 1972–7. doi:10.1016/j.cub.2008.11.021. PMC 2617768. PMID 19084405.
- ↑ Nadar VC, Lin S, Baas PW (April 2012). "Microtubule redistribution in growth cones elicited by focal inactivation of kinesin-5". J. Neurosci. 32 (17): 5783–94. doi:10.1523/JNEUROSCI.0144-12.2012. PMC 3347042. PMID 22539840.
- ↑ Liu M, Nadar VC, Kozielski F, Kozlowska M, Yu W, Baas PW (November 2010). "Kinesin-12, a mitotic microtubule-associated motor protein, impacts axonal growth, navigation, and branching". J. Neurosci. 30 (44): 14896–906. doi:10.1523/JNEUROSCI.3739-10.2010. PMC 3064264. PMID 21048148.
- ↑ Lin S, Liu M, Mozgova OI, Yu W, Baas PW (October 2012). "Mitotic motors coregulate microtubule patterns in axons and dendrites". J. Neurosci. 32 (40): 14033–49. doi:10.1523/JNEUROSCI.3070-12.2012. PMC 3482493. PMID 23035110.
- ↑ Yoon SY, Choi JE, Huh JW, Hwang O, Lee HS, Hong HN, Kim D (April 2005). "Monastrol, a selective inhibitor of the mitotic kinesin Eg5, induces a distinctive growth profile of dendrites and axons in primary cortical neuron cultures". Cell Motil. Cytoskeleton. 60 (4): 181–90. doi:10.1002/cm.20057. PMID 15751098.
- ↑ Lin S, Liu M, Son YJ, Timothy Himes B, Snow DM, Yu W, Baas PW (March 2011). "Inhibition of Kinesin-5, a microtubule-based motor protein, as a strategy for enhancing regeneration of adult axons". Traffic. 12 (3): 269–86. doi:10.1111/j.1600-0854.2010.01152.x. PMC 3037443. PMID 21166743.
- ↑ Rath O, Kozielski F (Aug 2012). "Kinesins and cancer". Nat Rev Cancer. 12 (8): 527–39. doi:10.1038/nrc3310. PMID 22825217. S2CID 20577157.
- 1 2 El-Nassan HB (2012). "Advances in the discovery of kinesin spindle protein (Eg5) inhibitors as antitumor agents". Eur J Med Chem. 62: 614–31. doi:10.1016/j.ejmech.2013.01.031. PMID 23434636.
- ↑ Sawin KE, Mitchison TJ (1995). "Mutations in the kinesin-like protein Eg5 disrupting localization to the mitotic spindle". Proc Natl Acad Sci U S A. 92 (10): 4289–93. Bibcode:1995PNAS...92.4289S. doi:10.1073/pnas.92.10.4289. PMC 41929. PMID 7753799.
- ↑ Rapley J, Nicolas M, Groen A, Regue L, Bertran MT, Caelles C, Avruch J, Roig J (2008). "The NIMA-family kinase Nek6 phosphorylates the kinesin Eg5 at a novel site necessary for mitotic spindle formation". J Cell Sci. 121 (Pt 23): 3912–21. doi:10.1242/jcs.035360. PMC 4066659. PMID 19001501.
- ↑ Liu M, Aneja R, Sun X, Xie S, Wang H, Wu X, Dong JT, Li M, Joshi HC, Zhou J (2008). "Parkin regulates Eg5 expression by Hsp70 ubiquitination-dependent inactivation of c-Jun NH2-terminal kinase". J Biol Chem. 283 (51): 35783–8. doi:10.1074/jbc.M806860200. PMID 18845538.
- ↑ Garcia K, Stumpff J, Duncan T, Su TT (2009). "Tyrosines in the kinesin-5 head domain are necessary for phosphorylation by Wee1 and for mitotic spindle integrity". Curr Biol. 19 (19): 1670–6. doi:10.1016/j.cub.2009.08.013. PMC 2762001. PMID 19800237.
- ↑ Eckerdt F, Eyers PA, Lewellyn AL, Prigent C, Maller JL (2008). "Spindle pole regulation by a discrete Eg5-interacting domain in TPX2". Curr Biol. 18 (7): 519–25. doi:10.1016/j.cub.2008.02.077. PMC 2408861. PMID 18372177.
- ↑ Ma N, Titus J, Gable A, Ross JL, Wadsworth P (2011). "TPX2 regulates the localization and activity of Eg5 in the mammalian mitotic spindle". J Cell Biol. 195 (1): 87–98. doi:10.1083/jcb.201106149. PMC 3187703. PMID 21969468.
- ↑ Blangy A, Arnaud L, Nigg EA (1997). "Phosphorylation by p34cdc2 protein kinase regulates binding of the kinesin-related motor HsEg5 to the dynactin subunit p150". J Biol Chem. 272 (31): 19418–24. doi:10.1074/jbc.272.31.19418. PMID 9235942.
- ↑ Iwakiri Y, Kamakura S, Hayase J, Sumimoto H (2013). "Interaction of NuMA protein with the kinesin Eg5: its possible role in bipolar spindle assembly and chromosome alignment". Biochem J. 451 (2): 195–204. doi:10.1042/BJ20121447. hdl:2324/1398274. PMID 23368718.
- ↑ Wilde A, Lizarraga SB, Zhang L, Wiese C, Gliksman NR, Walczak CE, Zheng Y (2001). "Ran stimulates spindle assembly by altering microtubule dynamics and the balance of motor activities". Nat Cell Biol. 3 (3): 221–7. doi:10.1038/35060000. PMID 11231570. S2CID 9536723.
- ↑ Koffa MD, Casanova CM, Santarella R, Kocher T, Wilm M, Mattaj IW (2006). "HURP is part of a Ran-dependent complex involved in spindle formation". Curr Biol. 16 (8): 743–54. doi:10.1016/j.cub.2006.03.056. PMID 16631581. S2CID 7466868.
- ↑ Maliga Z, Kapoor TM, Mitchison TJ (2002). "Evidence that monastrol is an allosteric inhibitor of the mitotic kinesin Eg5". Chem Biol. 9 (9): 989–96. doi:10.1016/S1074-5521(02)00212-0. PMID 12323373.
- ↑ Cochran JC, Krzysiak TC, Gilbert SP (2006). "Pathway of ATP hydrolysis by monomeric kinesin Eg5". Biochemistry. 45 (40): 12334–44. doi:10.1021/bi0608562. PMC 2288585. PMID 17014086.
- ↑ Vale RD (1996). "Switches, latches, and amplifiers: common themes of G proteins and molecular motors". J Cell Biol. 135 (2): 291–302. doi:10.1083/jcb.135.2.291. PMC 2121043. PMID 8896589.
- ↑ Kull FJ, Endow SA (2002). "Kinesin: switch I & II and the motor mechanism". J Cell Sci. 115 (Pt 1): 15–23. doi:10.1242/jcs.115.1.15. PMID 11801720.
- ↑ Parke CL, Wojcik EJ, Kim S, Worthylake DK (2010). "ATP hydrolysis in Eg5 kinesin involves a catalytic two-water mechanism". J Biol Chem. 285 (8): 5859–67. doi:10.1074/jbc.M109.071233. PMC 2820811. PMID 20018897.
- ↑ Jun B, Kim S (2010). "Real-time structural transitions are coupled to chemical steps in ATP hydrolysis by Eg5 kinesin". J Biol Chem. 285 (15): 11073–7. doi:10.1074/jbc.C110.103762. PMC 2856982. PMID 20154092.
- ↑ Chan Q, Nitta R, Inoue S, Hirokawa N (2013). "Structural Basis for the ATP-Induced Isomerization of Kinesin". J Mol Biol. 425 (11): 1869–80. doi:10.1016/j.jmb.2013.03.004. PMID 23500491.
- ↑ Onishi H, Mochizuki N, Morales MF (2004). "On the myosin catalysis of ATP hydrolysis". Biochemistry. 43 (13): 3757–63. doi:10.1021/bi040002m. PMID 15049682.
- ↑ Smith CA, Rayment I (1996). "X-ray structure of the magnesium(II).ADP.vanadate complex of the Dictyostelium discoideum myosin motor domain to 1.9 A resolution". Biochemistry. 35 (17): 5404–17. CiteSeerX 10.1.1.543.1030. doi:10.1021/bi952633+. PMID 8611530.
- ↑ Kapitein LC, Kwok BH, Weinger JS, Schmidt CF, Kapoor TM, Peterman EJ (2008). "Microtubule cross-linking triggers the directional motility of kinesin-5". J Cell Biol. 182 (3): 421–8. doi:10.1083/jcb.200801145. PMC 2500128. PMID 18678707.
- ↑ Kwok BH, Yang JG, Kapoor TM (2004). "The rate of bipolar spindle assembly depends on the microtubule-gliding velocity of the mitotic kinesin Eg5". Curr Biol. 14 (4): 1783–8. doi:10.1016/j.cub.2004.09.052. PMID 15458652.
- ↑ Weinger JS, Qiu M, Yang G, Kapoor TM (2011). "A nonmotor microtubule binding site in kinesin-5 is required for filament crosslinking and sliding". Curr Biol. 21 (2): 154–160. doi:10.1016/j.cub.2010.12.038. PMC 3049310. PMID 21236672.
- ↑ Valentine MT, Block SM (2009). "Force and premature binding of ADP can regulate the processivity of individual Eg5 dimers". Biophys J. 97 (6): 1671–7. Bibcode:2009BpJ....97.1671V. doi:10.1016/j.bpj.2009.07.013. PMC 2749793. PMID 19751672.
- ↑ Valentine MT, Fordyce PM, Krzysiak TC, Gilbert SP, Block SM (2006). "Individual dimers of the mitotic kinesin motor Eg5 step processively and support substantial loads in vitro". Nat Cell Biol. 8 (5): 470–6. doi:10.1038/ncb1394. PMC 1523314. PMID 16604065.
- ↑ Zhang Y, Xu W (August 2008). "Progress on kinesin spindle protein inhibitors as anti-cancer agents". Anticancer Agents Med Chem. 8 (6): 698–704. doi:10.2174/1871520610808060698. PMID 18690830.
- ↑ Gura, Trisha (21 September 2000). "A chemistry set for life". Nature International Weekly. 407 (6802): 282–284. doi:10.1038/35030189. PMID 11014160. S2CID 205008902. Retrieved 31 December 2012.
- ↑ Huszar D, Theoclitou ME, Skolnik J, Herbst R (2009). "Kinesin motor proteins as targets for cancer therapy". Cancer Metastasis Rev. 28 (1–2): 197–208. doi:10.1007/s10555-009-9185-8. PMID 19156502. S2CID 25682969.
- ↑ Compton DA (October 1999). "New tools for the antimitotic toolbox". Science. 286 (5441): 913–4. doi:10.1126/science.286.5441.913. PMID 10577242. S2CID 27786369.
- ↑ Hotha S, Yarrow JC, Yang JG, Garrett S, Renduchintala KV, Mayer TU, Kapoor TM (May 2003). "HR22C16: a potent small-molecule probe for the dynamics of cell division". Angew. Chem. Int. Ed. Engl. 42 (21): 2379–82. doi:10.1002/anie.200351173. PMID 12783501.
- ↑ Sakowicz R, Finer JT, Beraud C, Crompton A, Lewis E, Fritsch A, Lee Y, Mak J, Moody R, Turincio R, Chabala JC, Gonzales P, Roth S, Weitman S, Wood KW (May 2004). "Antitumor activity of a kinesin inhibitor". Cancer Res. 64 (9): 3276–80. doi:10.1158/0008-5472.can-03-3839. PMID 15126370.
- ↑ Turner J, Anderson R, Guo J, Beraud C, Fletterick R, Sakowicz R (July 2001). "Crystal structure of the mitotic spindle kinesin Eg5 reveals a novel conformation of the neck-linker". J. Biol. Chem. 276 (27): 25496–502. doi:10.1074/jbc.M100395200. PMID 11328809.
- ↑ Yan Y, Sardana V, Xu B, Homnick C, Halczenko W, Buser CA, Schaber M, Hartman GD, Huber HE, Kuo LC (2004). "Inhibition of a mitotic motor protein: where, how, and conformational consequences". J Mol Biol. 335 (2): 547–54. CiteSeerX 10.1.1.451.9558. doi:10.1016/j.jmb.2003.10.074. PMID 14672662.
- ↑ Learman SS, Kim CD, Stevens NS, Kim S, Wojcik EJ (2009). "NSC 622124 inhibits human Eg5 and other kinesins via interaction with the conserved microtubule-binding site". Biochemistry. 48 (8): 1754–62. doi:10.1021/bi801291q. PMC 3244877. PMID 19236100.
- ↑ Ulaganathan V, Talapatra SK, Rath O, Pannifer A, Hackney DD, Kozielski F (2013). "Structural insights into a unique inhibitor binding pocket in kinesin spindle protein". J Am Chem Soc. 135 (6): 2263–72. doi:10.1021/ja310377d. PMID 23305346.
- ↑ Cochran JC, Krzysiak TC, Gilbert SP (2005). "ATPase mechanism of Eg5 in the absence of microtubules: insight into microtubule activation and allosteric inhibition by monastrol". Biochemistry. 44 (50): 16633–48. doi:10.1021/bi051724w. PMC 2270472. PMID 16342954.
- ↑ Kwok BH, Kapitein LC, Kim JH, Peterman EJ, Schmidt CF, Kapoor TM (2006). "Allosteric inhibition of kinesin-5 modulates its processive directional motility". Nat Chem Biol. 2 (9): 480–5. doi:10.1038/nchembio812. PMID 16892050. S2CID 27535804.
- ↑ Crevel IM, Alonso MC, Cross RA (2004). "Monastrol stabilises an attached low-friction mode of Eg5". Curr Biol. 14 (11): R411–2. doi:10.1016/j.cub.2004.05.030. PMID 15182685. S2CID 15690493.
- ↑ Liu M, Aneja R, Liu C, Sun L, Gao J, Wang H, Dong JT, Sarli V, Giannis A, Joshi HC, Zhou J (2006). "Inhibition of the mitotic kinesin Eg5 up-regulates Hsp70 through the phosphatidylinositol 3-kinase/Akt pathway in multiple myeloma cells". J Biol Chem. 281 (26): 18090–7. doi:10.1074/jbc.M601324200. PMID 16627469.
- ↑ Orth JD, Tang Y, Shi J, Loy CT, Amendt C, Wilm C, Zenke FT, Mitchison TJ (2008). "Quantitative live imaging of cancer and normal cells treated with Kinesin-5 inhibitors indicates significant differences in phenotypic responses and cell fate". Mol Cancer Ther. 7 (11): 3480–9. doi:10.1158/1535-7163.MCT-08-0684. PMC 2597169. PMID 18974392.
- ↑ Miller K, Ng C, Ang P, Brufsky AM, Lee SC, Dees EC, Piccart M, Verrill M, Wardley A, Loftiss J, Bal J, Yeoh S, Hodge J, Williams D, Dar M, Ho PT. Phase II, open label study of SB-715992 (ispinesib) in subjects with advanced or metastatic breast cancer. 28th Annual San Antonio Breast Cancer Symposium.
- ↑ Deming D, Geiger P, Chen H, Kunnimalaiyaan M, Holen K (2010). "ZM336372 Induces Apoptosis Associated With Phosphorylation of GSK-3β in Pancreatic Adenocarcinoma Cell Lines". Cancer Chemother Pharmacol. 161 (1): 28–32. doi:10.1016/j.jss.2009.06.013. PMC 3379885. PMID 20031160.
- ↑ Cox CD, Breslin MJ, Mariano BJ, Coleman PJ, Buser CA, Walsh ES, Hamilton K, Huber HE, Kohl NE, Torrent M, Yan Y, Kuo LC, Hartman GD (2005). "Kinesin spindle protein (KSP) inhibitors. Part 1: The discovery of 3,5-diaryl-4,5-dihydropyrazoles as potent and selective inhibitors of the mitotic kinesin KSP". Bioorg Med Chem Lett. 15 (8): 2041–5. doi:10.1016/j.bmcl.2005.02.055. PMID 15808464.
- ↑ Kathman SJ, Williams DH, Hodge JP, Dar M (2007). "A Bayesian population PK-PD model of ispinesib-induced myelosuppression". Clin Pharmacol Ther. 81 (1): 88–94. doi:10.1038/sj.clpt.6100021. PMID 17186004. S2CID 34867346.
- ↑ Purcell JW, Davis J, Reddy M, Martin S, Samayoa K, Vo H, Thomsen K, Bean P, Kuo WL, Ziyad S, Billig J, Feiler HS, Gray JW, Wood KW, Cases S (2010). "Activity of the kinesin spindle protein inhibitor ispinesib (SB-715992) in models of breast cancer". Clin Cancer Res. 16 (2): 566–76. doi:10.1158/1078-0432.CCR-09-1498. PMC 2844774. PMID 20068098.
- ↑ Khoury HJ, Garcia-Manero G, Borthakur G, Kadia T, Foudray MC, Arellano M, Langston A, Bethelmie-Bryan B, Rush S, Litwiler K, Karan S, Simmons H, Marcus AI, Ptaszynski M, Kantarjian H (2012). "A phase 1 dose-escalation study of ARRY-520, a kinesin spindle protein inhibitor, in patients with advanced myeloid leukemias". Cancer. 118 (14): 3556–64. doi:10.1002/cncr.26664. PMC 4984525. PMID 22139909.
- ↑ Harrington TD, Naber N, Larson AG, Cooke R, Rice SE, Pate E (2011). "Analysis of the interaction of the Eg5 Loop5 with the nucleotide site". J Theor Biol. 289: 107–15. Bibcode:2011JThBi.289..107H. doi:10.1016/j.jtbi.2011.08.017. PMC 3191284. PMID 21872609.
- ↑ Behnke-Parks WM, Vendome J, Honig B, Maliga Z, Moores C, Rosenfeld SS (2011). "Loop L5 acts as a conformational latch in the mitotic kinesin Eg5". Journal of Biological Chemistry. 286 (7): 5242–53. doi:10.1074/jbc.M110.192930. PMC 3037637. PMID 21148480.
- ↑ Tcherniuk S, van Lis R, Kozielski F, Skoufias DA (March 2010). "Mutations in the human kinesin Eg5 that confer resistance to monastrol and S-trityl-L-cysteine in tumor derived cell lines" (PDF). Biochem. Pharmacol. 79 (6): 864–72. doi:10.1016/j.bcp.2009.11.001. PMID 19896928.
- ↑ Online Mendelian Inheritance in Man (OMIM): MCLMR - 152950
- ↑ Matthieu J Schlögel; Antonella Mendola; Elodie Fastré; Pradeep Vasudevan; Koen Devriendt; Thomy JL de Ravel; Hilde Van Esch; Ingele Casteels; Ignacio Arroyo Carrera; Francesca Cristofoli; Karen Fieggen; Katheryn Jones; Mark Lipson; Irina Balikova; Ami Singer; Maria Soller; María Mercedes Villanueva; Nicole Revencu; Laurence M Boon; Pascal Brouillard; Miikka Vikkula (May 2015). "No evidence of locus heterogeneity in familial microcephaly with or without chorioretinopathy, lymphedema, or mental retardation syndrome". Orphanet Journal of Rare Diseases. 10 (52): 52. doi:10.1186/s13023-015-0271-4. PMC 4464120. PMID 25934493.
Further reading
- Miki H, Setou M, Kaneshiro K, Hirokawa N (June 2001). "All kinesin superfamily protein, KIF, genes in mouse and human". Proc. Natl. Acad. Sci. U.S.A. 98 (13): 7004–11. Bibcode:2001PNAS...98.7004M. doi:10.1073/pnas.111145398. PMC 34614. PMID 11416179.
- Prince JA, Feuk L, Gu HF, Johansson B, Gatz M, Blennow K, Brookes AJ (November 2003). "Genetic variation in a haplotype block spanning IDE influences Alzheimer disease". Hum. Mutat. 22 (5): 363–71. doi:10.1002/humu.10282. PMID 14517947. S2CID 24508630.
- Yoon HG, Chan DW, Reynolds AB, Qin J, Wong J (September 2003). "N-CoR mediates DNA methylation-dependent repression through a methyl CpG binding protein Kaiso". Mol. Cell. 12 (3): 723–34. doi:10.1016/j.molcel.2003.08.008. PMID 14527417.
- Cassimeris L, Morabito J (April 2004). "TOGp, the human homolog of XMAP215/Dis1, is required for centrosome integrity, spindle pole organization, and bipolar spindle assembly". Mol. Biol. Cell. 15 (4): 1580–90. doi:10.1091/mbc.E03-07-0544. PMC 379257. PMID 14718566.
- Ertekin-Taner N, Allen M, Fadale D, Scanlin L, Younkin L, Petersen RC, Graff-Radford N, Younkin SG (April 2004). "Genetic variants in a haplotype block spanning IDE are significantly associated with plasma Abeta42 levels and risk for Alzheimer disease". Hum. Mutat. 23 (4): 334–42. doi:10.1002/humu.20016. PMID 15024728. S2CID 24885305.
- Tihy F, Kress M, Harper M, Dutrillaux B, Lemieux N (August 1992). "Localization of the human kinesin-related gene to band 10q24 by fluorescence in situ hybridization". Genomics. 13 (4): 1371–2. doi:10.1016/0888-7543(92)90075-4. PMID 1505978.
- Cochran JC, Sontag CA, Maliga Z, Kapoor TM, Correia JJ, Gilbert SP (September 2004). "Mechanistic analysis of the mitotic kinesin Eg5". J. Biol. Chem. 279 (37): 38861–70. doi:10.1074/jbc.M404203200. PMC 1356567. PMID 15247293.
- Cochran JC, Gatial JE, Kapoor TM, Gilbert SP (April 2005). "Monastrol inhibition of the mitotic kinesin Eg5". J. Biol. Chem. 280 (13): 12658–67. doi:10.1074/jbc.M413140200. PMC 1356610. PMID 15665380.
- Feuk L, McCarthy S, Andersson B, Prince JA, Brookes AJ (July 2005). "Mutation screening of a haplotype block around the insulin degrading enzyme gene and association with Alzheimer's disease". Am. J. Med. Genet. B Neuropsychiatr. Genet. 136B (1): 69–71. doi:10.1002/ajmg.b.30172. PMID 15858821. S2CID 20486238.
External links
- Baas, Peter. "Peter Baas Laboratory". Research Laboratory.
- Block, Steven
- Gilbert, Susan
- Kapoor, Tarun
- Kim, Sunyoung
- Kozielski, Frank
- Mitchison, Tim. "Tim Mitchison Laboratory". Mitchison Lab. Archived from the original on 2013-05-16. Retrieved 2012-12-31.
- Moores, Carolyn
- Rice, Sarah
- Rosenfeld, Steven
- Wadsworth, Pat. "Patricia Wadsworth Lab". Research Lab.
- Wojcik, Edward
- Worthylake, David
- Sharp, David. "David Sharp Lab". Research Lab.