| Medium spiny neuron | |
|---|---|
| Details | |
| Location | Basal ganglia |
| Shape | Spiny neuron |
| Function | Inhibitory projection neuron |
| Neurotransmitter | GABA |
| Presynaptic connections | Dopaminergic: VTA, SNc Glutamatergic: PFC, hippocampus, amygdala, thalamus, other |
| Postsynaptic connections | Other basal ganglia structures |
| Identifiers | |
| MeSH | D000094242 |
| NeuroLex ID | nifext_141 |
| Anatomical terms of neuroanatomy | |
Medium spiny neurons (MSNs), also known as spiny projection neurons (SPNs), are a special type of GABAergic inhibitory cell representing 95% of neurons within the human striatum, a basal ganglia structure.[1] Medium spiny neurons have two primary phenotypes (characteristic types): D1-type MSNs of the direct pathway and D2-type MSNs of the indirect pathway.[1][2][3] Most striatal MSNs contain only D1-type or D2-type dopamine receptors, but a subpopulation of MSNs exhibit both phenotypes.[1][2][3]
Direct pathway MSNs excite their ultimate basal ganglia output structure (such as the thalamus) and promote associated behaviors;[1] these neurons express D1-type dopamine receptors, adenosine A1 receptors, dynorphin peptides, and substance P peptides.[1][2] Indirect pathway MSNs inhibit their output structure and in turn inhibit associated behaviors;[1] these neurons express D2-type dopamine receptors, adenosine A2A receptors (A2A), DRD2–A2A heterotetramers, and enkephalin.[2][4] Both types express glutamate receptors (NMDAR and AMPAR), cholinergic receptors (M1 and M4)[5] and CB1 receptors are expressed on the somatodendritic area of both MSN types.[2][6] A subpopulation of MSNs contain both D1-type and D2-type receptors, with approximately 40% of striatal MSNs expressing both DRD1 and DRD2 mRNA.[1][2][3] In the nucleus accumbens (NAcc), these mixed-type MSNs that contain both D1-type and D2-type receptors are mostly contained in the NAcc shell.[1]
The dorsal striatal MSNs play a key role in initiating and controlling movements of the body, limbs, and eyes. The ventral striatal MSNs play a key role in motivation, reward, reinforcement, and aversion. Dorsal and ventral medium spiny neuron subtypes (i.e., direct D1-type and indirect D2-type) are identical phenotypes, but their output connections differ.[1][2]
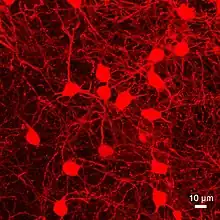
{kind=link}
Appearance and location
The medium spiny neurons are medium-sized neurons (~15 microns in diameter, ~12–13 microns in the mouse) with large and extensive dendritic trees (~500 microns in diameter).[8] Striatal direct pathway MSNs (dMSNs) project directly to the globus pallidus internal (GPi) and substantia nigra pars reticulata (SNpr) whereas striatal indirect pathway MSNs (iMSNs) ultimately project to these two structures via an intermediate connection to the globus pallidus external (GPe) and ventral pallidum (VP).[1] The GPe and VP send a GABAergic projection to the subthalamic nucleus, which then sends glutamatergic projections to the GPi and SNpr.[1] Both the GPi and SNpr send inhibitory projections to nuclei within the thalamus.[1]
Function
MSNs are inhibitory GABAergic neurons, but the effect of direct MSNs (dMSNs) and indirect MSNs (iMSNs) on their ultimate output structures differs: dMSNs excite, while iMSNs inhibit, their basal ganglia output structures (e.g., the thalamus).[1] Within the basal ganglia, there are several complex circuits of neuronal loops all of which include medium spiny neurons.
The cortical, thalamic, and brain-stem inputs that arrive at the medium spiny neurons show a vast divergence in that each incoming axon forms contacts with many spiny neurons and each spiny neuron receives a vast amount of input from different incoming axons. Since these inputs are glutamatergic they exhibit an excitatory influence on the inhibitory medium spiny neurons.
There are also interneurons in the striatum which regulate the excitability of the medium spiny neurons. The synaptic connections between a particular GABAergic interneuron, the parvalbumin expressing fast-spiking interneuron, and spiny neurons are close to the spiny neurons' soma, or cell body.[9] Recall that excitatory postsynaptic potentials caused by glutamatergic inputs at the dendrites of the spiny neurons only cause an action potential when the depolarization wave is strong enough upon entering the cell soma. Since the fast-spiking interneurons influence is located so closely to this critical gate between the dendrites and the soma, they can readily regulate the generation of an action potential. Additionally, other types of GABAergic interneurons make connections with the spiny neurons. These include interneurons that express tyrosine hydroxylase[10][11] and neuropeptide Y.[12][13]
Dorsal striatal MSNs
Direct pathway
Anatomy
The direct pathway within the basal ganglia receives excitatory input from the cortex, thalamus, and other brain regions. In the direct pathway, medium spiny neurons project to the internal division of the globus pallidus (GPi) or the substantia nigra pars reticula (SNpr or SNr). These nuclei project to the deep layer of the superior colliculus and control fast eye movements (saccades),[14] and also project to the ventral thalamus, which in turn projects to upper motor neurons in the primary motor cortex (precentral gyrus).[15] The SNr and GPi outputs are both tonically active inhibitory nuclei and are thus constantly inhibiting the thalamus (and thus motor cortex). However, transient activity in (inhibitory) direct pathway medium spiny neurons ultimately disinhibits thalamus projections to the motor cortex and enables movement.[16]
Indirect pathway
Anatomy
The indirect pathway also receives excitatory input from various brain regions. Indirect pathway medium spiny neurons project to the external segment of the globus pallidus (GPe). Like the GPi, the GPe is a tonically active inhibitory nucleus. The GPe projects to the excitatory subthalamic nucleus (STN), which in turn projects to the GPi and SNr.[15] When the indirect pathway is not activated, activity in the STN is suppressed by the GPe, which translates to decreased SNr/GPi activity downstream and thus increased thalamic and motor cortex neuron activity. When indirect pathway neurons fire, GPe neurons are inhibited, which disinhibits the STN. The STN then excites SNr/GPi neurons, suppressing thalamus/motor cortex activity.[16]
Functional distinctions
Classic models of striatal function have posited that activation of the direct pathway leads to movement, whereas activation of the indirect pathway leads to the termination of movement.[17][18] This model is supported by experiments demonstrating that optogenetically stimulating direct pathway medium spiny neurons increases locomotion, whereas stimulating indirect pathway medium spiny neurons inhibits locomotion.[19] The balance of direct/indirect activity in movement is supported by evidence from neurodegenerative disorders, including Parkinson's disease (PD), which is characterized by loss of dopamine neurons projecting to the striatum, hypoactivity in direct pathway and hyperactivity in indirect pathway neurons, along with motor dysfunction.[20] This results in loss of normal action selection, as loss of dopamine drives activity in the indirect pathway, globally inhibiting all motor paradigms. This may explain impaired action initiation, slowed actions (bradykinesia), and impaired voluntary motor initiation in Parkinson's patients. On the other hand, Huntington's disease, which is characterized by preferential degradation of indirect pathway medium spiny neurons, results in unwanted movements (chorea) that may result from impaired movement inhibition and predominant direct pathway activity.[21] An alternative related hypothesis is that the striatum controls action initiation and selection via a ’center-surround’ architecture, where activation of a subset of direct pathway neurons initiates movements while closely related motor patterns represented by surrounding neurons are inhibited by lateral inhibition via indirect pathway neurons.[22] This specific hypothesis is supported by recent calcium-imaging work showing that direct and indirect pathway medium spiny neurons encoding specific actions are located in spatially organized ensembles.[23]
Despite the abundance of evidence for the initiation/termination model, recent evidence using transgenic mice expressing calcium indicators in either the direct or indirect pathway demonstrated that both pathways are active at action initiation, but neither are active during inactivity,[24] a finding which has been replicated using simultaneous two-channel calcium imaging.[25] This has led to somewhat of a paradigm shift in models of striatal functioning, such that newer models posit that the direct pathway facilitates wanted movements, whereas the indirect pathway simultaneously inhibits unwanted movements.[26][27] Indeed, more sophisticated techniques and analyses, such as state-dependent optogenetics, have revealed that both pathways are heavily involved in action sequence execution,[28] and that specifically, both striatal pathways are involved in element-level action control.[29] However, direct pathway medium spiny neurons mostly signal sequence initiation/termination and indirect pathway medium spiny neurons may signal switching between subsequences of a given action sequence.[30] Other evidence suggests that the direct and indirect pathway oppositely influence the termination of movement—specifically, the relative timing of their activity determines if an action will be terminated.[31]
Recent experiments have established that the direct and indirect pathways of the dorsal striatum are not solely involved in movement. Initial experiments in an intracranial self-stimulation paradigm suggested opposing roles in reinforcement for the two pathways; specifically, stimulation of direct pathway medium spiny neurons was found to be reinforcing, whereas stimulation of indirect pathway medium spiny neurons was aversive.[32] However, a subsequent study (using more physiologically relevant stimulation parameters) found that direct and indirect pathway stimulation was reinforcing, but that pathway-specific stimulation resulted in the development of different action strategies.[33] Regardless, these studies suggest a critical role for reinforcement in the dorsal striatum, as opposed to the striatum only serving a role in movement control.
Ventral striatal MSNs
Direct pathway
The direct pathway of the ventral striatum within the basal ganglia mediates reward-based learning and appetitive incentive salience, which is assigned to rewarding stimuli.[34]
Indirect pathway
The indirect pathway of the ventral striatum within the basal ganglia mediates aversion-based learning and aversive motivational salience, which is assigned to aversive stimuli.[34]
See also
References
- 1 2 3 4 5 6 7 8 9 10 11 12 13 Yager LM, Garcia AF, Wunsch AM, Ferguson SM (August 2015). "The ins and outs of the striatum: Role in drug addiction". Neuroscience. 301: 529–541. doi:10.1016/j.neuroscience.2015.06.033. PMC 4523218. PMID 26116518.
[The striatum] receives dopaminergic inputs from the ventral tegmental area (VTA) and the substantia nigra (SNr) and glutamatergic inputs from several areas, including the cortex, hippocampus, amygdala, and thalamus (Swanson, 1982; Phillipson and Griffiths, 1985; Finch, 1996; Groenewegen et al., 1999; Britt et al., 2012). These glutamatergic inputs make contact on the heads of dendritic spines of the striatal GABAergic medium spiny projection neurons (MSNs) whereas dopaminergic inputs synapse onto the spine neck, allowing for an important and complex interaction between these two inputs in modulation of MSN activity ... It should also be noted that there is a small population of neurons in the NAc that coexpress both D1 and D2 receptors, though this is largely restricted to the NAc shell (Bertran- Gonzalez et al., 2008). ... Neurons in the NAc core and NAc shell subdivisions also differ functionally. The NAc core is involved in the processing of conditioned stimuli whereas the NAc shell is more important in the processing of unconditioned stimuli; Classically, these two striatal MSN populations are thought to have opposing effects on basal ganglia output. Activation of the dMSNs causes a net excitation of the thalamus resulting in a positive cortical feedback loop; thereby acting as a 'go' signal to initiate behavior. Activation of the iMSNs, however, causes a net inhibition of thalamic activity resulting in a negative cortical feedback loop and therefore serves as a 'brake' to inhibit behavior ... there is also mounting evidence that iMSNs play a role in motivation and addiction (Lobo and Nestler, 2011; Grueter et al., 2013). For example, optogenetic activation of NAc core and shell iMSNs suppressed the development of a cocaine CPP whereas selective ablation of NAc core and shell iMSNs ... enhanced the development and the persistence of an amphetamine CPP (Durieux et al., 2009; Lobo et al., 2010). These findings suggest that iMSNs can bidirectionally modulate drug reward. ... Together these data suggest that iMSNs normally act to restrain drug-taking behavior and recruitment of these neurons may in fact be protective against the development of compulsive drug use.
- 1 2 3 4 5 6 7 Ferré S, Lluís C, Justinova Z, Quiroz C, Orru M, Navarro G, Canela EI, Franco R, Goldberg SR (June 2010). "Adenosine-cannabinoid receptor interactions. Implications for striatal function". Br. J. Pharmacol. 160 (3): 443–453. doi:10.1111/j.1476-5381.2010.00723.x. PMC 2931547. PMID 20590556.
Two classes of MSNs, which are homogeneously distributed in the striatum, can be differentiated by their output connectivity and their expression of dopamine and adenosine receptors and neuropeptides. In the dorsal striatum (mostly represented by the nucleus caudate-putamen), enkephalinergic MSNs connect the striatum with the globus pallidus (lateral globus pallidus) and express the peptide enkephalin and a high density of dopamine D2 and adenosine A2A receptors (they also express adenosine A1 receptors), while dynorphinergic MSNs connect the striatum with the substantia nigra (pars compacta and reticulata) and the entopeduncular nucleus (medial globus pallidus) and express the peptides dynorphin and substance P and dopamine D1 and adenosine A1 but not A2A receptors (Ferréet al., 1997; Gerfen, 2004; Quiroz et al., 2009). These two different phenotypes of MSN are also present in the ventral striatum (mostly represented by the nucleus accumbens and the olfactory tubercle). However, although they are phenotypically equal to their dorsal counterparts, they have some differences in terms of connectivity. First, not only enkephalinergic but also dynorphinergic MSNs project to the ventral counterpart of the lateral globus pallidus, the ventral pallidum, which, in fact, has characteristics of both the lateral and medial globus pallidus in its afferent and efferent connectivity. In addition to the ventral pallidum, the medial globus pallidus and the substantia nigra-VTA, the ventral striatum sends projections to the extended amygdala, the lateral hypothalamus and the pedunculopontine tegmental nucleus. Finally, unlike the dorsal striatum, the substantia nigra pars reticulata is not a main target area for the ventral striatum, which preferentially directs its midbrain output to the substantia nigra pars compacta and the VTA (Heimer et al., 1995; Robertson and Jian, 1995; Ferré, 1997). It is also important to mention that a small percentage of MSNs have a mixed phenotype and express both D1 and D2 receptors (Surmeier et al., 1996). ... A2A receptors are localized predominantly postsynaptically in the dendritic spine of enkephalinergic but not dynorphinergic MSNs, co-localized with D2 receptors ... Presynaptically, CB1 receptors are localized in GABAergic terminals of interneurons or collaterals from MSNs, and also in glutamatergic but not in dopaminergic terminals ... Postsynaptically, CB1 receptors are localized in the somatodendritic area of MSN (Rodriguez et al., 2001; Pickel et al., 2004; 2006; Köfalvi et al., 2005) and both enkephalinergic and dynorphinergic MSNs express CB1 receptors (Martín et al., 2008).
- 1 2 3 Nishi A, Kuroiwa M, Shuto T (July 2011). "Mechanisms for the modulation of dopamine d(1) receptor signaling in striatal neurons". Front Neuroanat. 5: 43. doi:10.3389/fnana.2011.00043. PMC 3140648. PMID 21811441.
Dopamine plays critical roles in the regulation of psychomotor functions in the brain (Bromberg-Martin et al., 2010; Cools, 2011; Gerfen and Surmeier, 2011). The dopamine receptors are a superfamily of heptahelical G protein-coupled receptors, and are grouped into two categories, D1-like (D1, D5) and D2-like (D2, D3, D4) receptors, based on functional properties to stimulate adenylyl cyclase (AC) via Gs/olf and to inhibit AC via Gi/o, respectively ... It has been demonstrated that D1 receptors form the hetero-oligomer with D2 receptors, and that the D1–D2 receptor hetero-oligomer preferentially couples to Gq/PLC signaling (Rashid et al., 2007a,b). The expression of dopamine D1 and D2 receptors are largely segregated in direct and indirect pathway neurons in the dorsal striatum, respectively (Gerfen et al., 1990; Hersch et al., 1995; Heiman et al., 2008). However, some proportion of medium spiny neurons are known to expresses both D1 and D2 receptors (Hersch et al., 1995). Gene expression analysis using single cell RT-PCR technique estimated that 40% of medium spiny neurons express both D1 and D2 receptor mRNA (Surmeier et al., 1996).
- ↑ Ferré S, Bonaventura J, Tomasi D, Navarro G, Moreno E, Cortés A, Lluís C, Casadó V, Volkow ND (June 2015). "Allosteric mechanisms within the adenosine A2A-dopamine D2 receptor heterotetramer". Neuropharmacology. 104: 154–60. doi:10.1016/j.neuropharm.2015.05.028. PMC 5754196. PMID 26051403.
caffeine-induced increases in D2R availability in the ventral striatum were associated with caffeine-induced increases in alertness (Volkow et al., 2015). ... Fig. 2. Brain maps showing significant differences in D2R/D3R availability (nondisplaceable binding potential or BPND), between placebo and caffeine
- ↑ Benarroch EE (July 2012). "Effects of acetylcholine in the striatum. Recent insights and therapeutic implications". Neurology. 79 (3): 274–81. doi:10.1212/WNL.0b013e31825fe154. PMID 22802594. S2CID 29003596.
- ↑ Gardoni F, Bellone C (2015). "Modulation of the glutamatergic transmission by Dopamine: a focus on Parkinson, Huntington and Addiction diseases". Frontiers in Cellular Neuroscience. 9: 25. doi:10.3389/fncel.2015.00025. PMC 4345909. PMID 25784855.
In particular in the striatum the release of DA rapidly influences synaptic transmission modulating both AMPA and NMDA receptors.
- ↑ Reinius B, Blunder M, Brett FM, Eriksson A, Patra K, Jonsson J, Jazin E, Kullander K (27 March 2015). "Conditional targeting of medium spiny neurons in the striatal matrix". Frontiers in Behavioral Neuroscience. 9: 71. doi:10.3389/fnbeh.2015.00071. PMC 4375991. PMID 25870547.
- ↑ Kawaguchi Y, Wilson CJ, Emson PC (October 1990). "Projection subtypes of rat neostriatal matrix cells revealed by intracellular injection of biocytin". The Journal of Neuroscience. 10 (10): 3421–38. doi:10.1523/JNEUROSCI.10-10-03421.1990. PMC 6570194. PMID 1698947.
- ↑ Tepper JM, Wilson CJ, Koós T (August 2008). "Feedforward and feedback inhibition in neostriatal GABAergic spiny neurons". Brain Research Reviews. 58 (2): 272–81. doi:10.1016/j.brainresrev.2007.10.008. PMC 2562631. PMID 18054796.
- ↑ Ibáñez-Sandoval O, Tecuapetla F, Unal B, Shah F, Koós T, Tepper JM (May 2010). "Electrophysiological and morphological characteristics and synaptic connectivity of tyrosine hydroxylase-expressing neurons in adult mouse striatum". The Journal of Neuroscience. 30 (20): 6999–7016. doi:10.1523/JNEUROSCI.5996-09.2010. PMC 4447206. PMID 20484642.
- ↑ Tepper JM, Tecuapetla F, Koós T, Ibáñez-Sandoval O (December 2010). "Heterogeneity and diversity of striatal GABAergic interneurons". Frontiers in Neuroanatomy. 4: 150. doi:10.3389/fnana.2010.00150. PMC 3016690. PMID 21228905.
- ↑ English DF, Ibanez-Sandoval O, Stark E, Tecuapetla F, Buzsáki G, Deisseroth K, Tepper JM, Koos T (December 2011). "GABAergic circuits mediate the reinforcement-related signals of striatal cholinergic interneurons". Nature Neuroscience. 15 (1): 123–30. doi:10.1038/nn.2984. PMC 3245803. PMID 22158514.
- ↑ Ibáñez-Sandoval O, Tecuapetla F, Unal B, Shah F, Koós T, Tepper JM (November 2011). "A novel functionally distinct subtype of striatal neuropeptide Y interneuron". The Journal of Neuroscience. 31 (46): 16757–69. doi:10.1523/JNEUROSCI.2628-11.2011. PMC 3236391. PMID 22090502.
- ↑ Hikosaka O, Takikawa Y, Kawagoe R (July 2000). "Role of the basal ganglia in the control of purposive saccadic eye movements". Physiological Reviews. 80 (3): 953–78. doi:10.1152/physrev.2000.80.3.953. PMID 10893428.
- 1 2 Lanciego JL, Luquin N, Obeso JA (December 2012). "Functional neuroanatomy of the basal ganglia". Cold Spring Harbor Perspectives in Medicine. 2 (12): a009621. doi:10.1101/cshperspect.a009621. PMC 3543080. PMID 23071379.
- 1 2 Freeze BS, Kravitz AV, Hammack N, Berke JD, Kreitzer AC (November 2013). "Control of basal ganglia output by direct and indirect pathway projection neurons". The Journal of Neuroscience. 33 (47): 18531–9. doi:10.1523/JNEUROSCI.1278-13.2013. PMC 3834057. PMID 24259575.
- ↑ Albin RL, Young AB, Penney JB (October 1989). "The functional anatomy of basal ganglia disorders" (PDF). Trends in Neurosciences. 12 (10): 366–75. doi:10.1016/0166-2236(89)90074-x. hdl:2027.42/28186. PMID 2479133. S2CID 8112392.
- ↑ DeLong MR (July 1990). "Primate models of movement disorders of basal ganglia origin". Trends in Neurosciences. 13 (7): 281–5. doi:10.1016/0166-2236(90)90110-v. PMID 1695404. S2CID 4057704.
- ↑ Kravitz AV, Freeze BS, Parker PR, Kay K, Thwin MT, Deisseroth K, Kreitzer AC (July 2010). "Regulation of parkinsonian motor behaviours by optogenetic control of basal ganglia circuitry". Nature. 466 (7306): 622–6. Bibcode:2010Natur.466..622K. doi:10.1038/nature09159. PMC 3552484. PMID 20613723.
- ↑ Bezard E, Jaber M, Gonon F, Boireau A, Bloch B, Gross CE (August 2000). "Adaptive changes in the nigrostriatal pathway in response to increased 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced neurodegeneration in the mouse". The European Journal of Neuroscience. 12 (8): 2892–900. doi:10.1046/j.1460-9568.2000.00180.x. PMID 10971632. S2CID 8984006.
- ↑ Roze E, Bonnet C, Betuing S, Caboche J (2010). "Huntington's Disease". Diseases of DNA Repair. Advances in Experimental Medicine and Biology. Vol. 685. pp. 45–63. doi:10.1007/978-1-4419-6448-9_5. ISBN 978-1-4419-6447-2. PMID 20687494.
- ↑ Schroll H, Hamker FH (December 2013). "Computational models of basal-ganglia pathway functions: focus on functional neuroanatomy". Frontiers in Systems Neuroscience. 7: 122. doi:10.3389/fnsys.2013.00122. PMC 3874581. PMID 24416002.
- ↑ Klaus A, Martins GJ, Paixao VB, Zhou P, Paninski L, Costa RM (August 2017). "The Spatiotemporal Organization of the Striatum Encodes Action Space". Neuron. 95 (5): 1171–1180.e7. doi:10.1016/j.neuron.2017.08.015. PMC 5584673. PMID 28858619.
- ↑ Cui G, Jun SB, Jin X, Pham MD, Vogel SS, Lovinger DM, Costa RM (February 2013). "Concurrent activation of striatal direct and indirect pathways during action initiation". Nature. 494 (7436): 238–42. Bibcode:2013Natur.494..238C. doi:10.1038/nature11846. PMC 4039389. PMID 23354054.
- ↑ Markowitz JE, Gillis WF, Beron CC, Neufeld SQ, Robertson K, Bhagat ND, Peterson RE, Peterson E, Hyun M, Linderman SW, Sabatini BL, Datta SR (June 2018). "The Striatum Organizes 3D Behavior via Moment-to-Moment Action Selection". Cell. 174 (1): 44–58.e17. doi:10.1016/j.cell.2018.04.019. PMC 6026065. PMID 29779950.
- ↑ Nambu A (December 2008). "Seven problems on the basal ganglia". Current Opinion in Neurobiology. 18 (6): 595–604. doi:10.1016/j.conb.2008.11.001. PMID 19081243. S2CID 24956799.
- ↑ Calabresi P, Picconi B, Tozzi A, Ghiglieri V, Di Filippo M (August 2014). "Direct and indirect pathways of basal ganglia: a critical reappraisal". Nature Neuroscience. 17 (8): 1022–30. doi:10.1038/nn.3743. PMID 25065439. S2CID 8983260.
- ↑ Tecuapetla F, Jin X, Lima SQ, Costa RM (July 2016). "Complementary Contributions of Striatal Projection Pathways to Action Initiation and Execution". Cell. 166 (3): 703–715. doi:10.1016/j.cell.2016.06.032. PMID 27453468.
- ↑ Jin X, Tecuapetla F, Costa RM (March 2014). "Basal ganglia subcircuits distinctively encode the parsing and concatenation of action sequences". Nature Neuroscience. 17 (3): 423–30. doi:10.1038/nn.3632. PMC 3955116. PMID 24464039.
- ↑ Geddes CE, Li H, Jin X (June 2018). "Optogenetic Editing Reveals the Hierarchical Organization of Learned Action Sequences". Cell. 174 (1): 32–43.e15. doi:10.1016/j.cell.2018.06.012. PMC 6056013. PMID 29958111.
- ↑ Atallah HE, McCool AD, Howe MW, Graybiel AM (June 2014). "Neurons in the ventral striatum exhibit cell-type-specific representations of outcome during learning". Neuron. 82 (5): 1145–56. doi:10.1016/j.neuron.2014.04.021. PMC 4108162. PMID 24908491.
- ↑ Kravitz AV, Tye LD, Kreitzer AC (June 2012). "Distinct roles for direct and indirect pathway striatal neurons in reinforcement". Nature Neuroscience. 15 (6): 816–8. doi:10.1038/nn.3100. PMC 3410042. PMID 22544310.
- ↑ Vicente AM, Galvão-Ferreira P, Tecuapetla F, Costa RM (April 2016). "Direct and indirect dorsolateral striatum pathways reinforce different action strategies". Current Biology. 26 (7): R267-9. doi:10.1016/j.cub.2016.02.036. PMC 4826435. PMID 27046807.
- 1 2 Baliki MN, Mansour A, Baria AT, Huang L, Berger SE, Fields HL, Apkarian AV (October 2013). "Parceling human accumbens into putative core and shell dissociates encoding of values for reward and pain". The Journal of Neuroscience. 33 (41): 16383–93. doi:10.1523/JNEUROSCI.1731-13.2013. PMC 3792469. PMID 24107968.
Recent evidence indicates that inactivation of D2 receptors, in the indirect striatopallidal pathway in rodents, is necessary for both acquisition and expression of aversive behavior, and direct pathway D1 receptor activation controls reward-based learning (Hikida et al., 2010; Hikida et al., 2013). It seems we can conclude that direct and indirect pathways of the NAc, via D1 and D2 receptors, subserve distinct anticipation and valuation roles in the shell and core of NAc, which is consistent with observations regarding spatial segregation and diversity of responses of midbrain dopaminergic neurons for rewarding and aversive conditions, some encoding motivational value, others motivational salience, each connected with distinct brain networks and having distinct roles in motivational control (Bromberg-Martin et al., 2010; Cohen et al., 2012; Lammel et al., 2013). ... Thus, the previous results, coupled with the current observations, imply that the NAc pshell response reflects a prediction/anticipation or salience signal, and the NAc pcore response is a valuation response (reward predictive signal) that signals the negative reinforcement value of cessation of pain (i.e., anticipated analgesia).
Further reading
- Bear, Mark F; Connors, Barry W.; Paradiso, Michael A., Neuroscience, Exploring the Brain, Lippincott Williams & Wilkins; Third Edition (1 February 2006). ISBN 0-7817-6003-8
- Kandel, E. (2006). Principles of neuroscience. (5th Ed.) Wadsworth
- Purves, D., Augustine, G.J. & Fitzpatrick, D. (2004). Neuroscience. (3rd Ed.). SInauer Associates
- Cell Centered Database – Medium spiny neuron