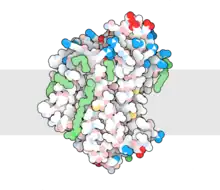
| NRAMP family | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||
| Identifiers | |||||||||||
| Symbol | ? | ||||||||||
| Pfam | PF01566 | ||||||||||
| TCDB | 2.A.55 | ||||||||||
| OPM superfamily | 64 | ||||||||||
| OPM protein | 4wgv | ||||||||||
| |||||||||||
Natural resistance-associated macrophage proteins (Nramps), also known as metal ion (Mn2+-iron) transporters (TC# 2.A.55), are a family of metal transport proteins found throughout all domains of life. Taking on an eleven-helix LeuT fold, the Nramp family is a member of the large APC Superfamily of secondary carriers.[1] They transport a variety of transition metals such as manganese, cadmium, and manganese using an alternating access mechanism characteristic of secondary transporters.[2]
The name "natural resistance-associated" macrophage proteins arises from the role in resistance of intracellular bacterial pathogens played by some animal homologs. Several human pathologies may result from defects in Nramp-dependent Fe2+ or Mn2+ transport, including iron overload, neurodegenerative diseases and innate susceptibility to infectious diseases.[3]
Human homologs
Humans and rodents possess two distinct Nramp proteins. The broad specificity NRAMP2 (DMT1) transports a range of divalent metal cations. Studies have shown that it transports Fe2+ and H+ with a 1:1 stoichiometry and apparent affinities of 6 μm and about 1 μm, respectively. Variable H+:Fe2+ stoichiometry has also been reported. The order of substrate preference for NRAMP2 is:
Fe2+> Zn2+> Mn2+> Co2+> Ca2+> Cu2+> Ni2+> Pb2+
Many of these ions can inhibit iron absorption. Mutation of NRAMP2 in rodents leads to defective endosomal iron export within the ferritin cycle, impaired intestinal iron absorption and microcytic anemia. Symptoms of Mn2+ deficiency are also seen. It is found in apical membranes of intestinal epithelial cells but also in late endosomes and lysosomes.
In contrast to the widely expressed NRAMP2, NRAMP1 is expressed primarily in macrophages and monocytes and appears to have a preference for Mn2+ rather than Fe2+. NRAMP1 (TC# 2.A.55.2.3) has been reported to function by metal:H+ antiport.[4] It is hypothesized that a deficiency for Mn2+ or some other metal prevents the generation of reactive oxygenic and nitrogenic compounds that are used by macrophage to combat pathogens. This hypothesis is supported by studies on the bacterial Nramp homologs which exhibit extremely high selectivity for Mn2+ over Fe2+, Zn2+ and other divalent cations.[5] Regulation of these transporters in bacteria can occur through Fur, OxyR, and most commonly a DtxR homolog, MntR.
Smf and other homologues
The Smf1 protein of Saccharomyces cerevisiae appears to catalyze high-affinity (KM = 0.3 μm) Mn2+ uptake while the closely related Smf2 protein may catalyze low affinity (KM = 60 μm) Mn2+ uptake in the same organism. Both proteins also mediate H+-dependent Fe2+ uptake. These proteins are of 575 and 549 amino acyl residues in length and are predicted to have 8-12 transmembrane α-helical spanners. The E. coli homologue of 412 aas exhibits 11 putative and confirmed TMSs with the N-terminus in and the C-terminus out. The yeast proteins may be localized to the vacuole and/or the plasma membrane of the yeast cell. Indirect and some direct experiments suggest that they may be able to transport several heavy metals including Mn2+, Cu2+, Cd2+ and Co2+. A third yeast protein, Smf3p, appears to be exclusively intracellular, possibly in the Golgi. NRAMP2 (Slc11A2) of Homo sapiens (TC# 2.A.55.2.1) has a 12 TMS topology with intracellular N- and C-termini. Two-fold structural symmetry in the arrangement of membrane helices for TM1-5 and TM6-10 (conserved Slc2 hydrophobic core) is suggested.[6]
Transport reaction
The generalized transport reaction catalyzed by NRAMP family proteins is:
Me2+ (out) + H+ (out) ⇌ Me2+ (in) + H+ (in).
Structure and Mechanism
All Nramp proteins have eleven to twelve transmembrane helices, the first ten of which form a canonical LeuT fold, common throughout the APC superfamily. Metal uptake in Nramp proteins is typically stimulated by acidic pH and accompanied by proton influx,[7] although many homologs have also shown proton uniport.[8] This has been explained in the Deinococcus radiodurans homolog as the result of spatially segregated metal and proton pathways that rely on a longer-range allosteric connection rather than the direct structural connection seen in canonical symporters.[9] Metal uptake requires alternating access bulk conformation change, in which the protein changes from an outward-open state to an inward-open state upon metal binding, while proton uptake can occur through a simpler channel-like mechanism.[2]
See also
References
- ↑ Vastermark A, Wollwage S, Houle ME, Rio R, Saier MH (October 2014). "Expansion of the APC superfamily of secondary carriers". Proteins. 82 (10): 2797–811. doi:10.1002/prot.24643. PMC 4177346. PMID 25043943.
- 1 2 Bozzi, Aaron T; Zimanyi, Christina M; Nicoludis, John M; Lee, Brandon K; Zhang, Casey H; Gaudet, Rachelle (2019-02-04). Faraldo-Gómez, José D; Marletta, Michael A; Zhou, Ming; Slotboom, Dirk; Rudnick, Gary (eds.). "Structures in multiple conformations reveal distinct transition metal and proton pathways in an Nramp transporter". eLife. 8: e41124. doi:10.7554/eLife.41124. ISSN 2050-084X. PMC 6398981. PMID 30714568.
- ↑ Cellier MF (2012-01-01). "Nramp: from sequence to structure and mechanism of divalent metal import". Current Topics in Membranes. 69: 249–93. doi:10.1016/B978-0-12-394390-3.00010-0. PMID 23046654.
- ↑ Techau ME, Valdez-Taubas J, Popoff JF, Francis R, Seaman M, Blackwell JM (December 2007). "Evolution of differences in transport function in Slc11a family members". The Journal of Biological Chemistry. 282 (49): 35646–56. doi:10.1074/jbc.M707057200. PMID 17932044.
- ↑ Wei W, Chai T, Zhang Y, Han L, Xu J, Guan Z (January 2009). "The Thlaspi caerulescens NRAMP homologue TcNRAMP3 is capable of divalent cation transport". Molecular Biotechnology. 41 (1): 15–21. doi:10.1007/s12033-008-9088-x. PMID 18663607. S2CID 5455501.
- ↑ Czachorowski M, Lam-Yuk-Tseung S, Cellier M, Gros P (September 2009). "Transmembrane topology of the mammalian Slc11a2 iron transporter". Biochemistry. 48 (35): 8422–34. doi:10.1021/bi900606y. PMC 2736113. PMID 19621945.
- ↑ Ehrnstorfer, Ines A.; Manatschal, Cristina; Arnold, Fabian M.; Laederach, Juerg; Dutzler, Raimund (2017-01-06). "Structural and mechanistic basis of proton-coupled metal ion transport in the SLC11/NRAMP family". Nature Communications. 8 (1): 14033. Bibcode:2017NatCo...814033E. doi:10.1038/ncomms14033. ISSN 2041-1723. PMC 5230734. PMID 28059071.
- ↑ Gunshin, H.; Mackenzie, B.; Berger, U. V.; Gunshin, Y.; Romero, M. F.; Boron, W. F.; Nussberger, S.; Gollan, J. L.; Hediger, M. A. (1997-07-31). "Cloning and characterization of a mammalian proton-coupled metal-ion transporter". Nature. 388 (6641): 482–488. Bibcode:1997Natur.388..482G. doi:10.1038/41343. ISSN 0028-0836. PMID 9242408. S2CID 4416482.
- ↑ Bozzi, Aaron T.; Bane, Lukas B.; Zimanyi, Christina M.; Gaudet, Rachelle (2019-12-02). "Unique structural features in an Nramp metal transporter impart substrate-specific proton cotransport and a kinetic bias to favor import". Journal of General Physiology. 151 (12): 1413–1429. doi:10.1085/jgp.201912428. ISSN 0022-1295. PMC 6888756. PMID 31619456.
As of 2 February 2016, this article is derived in whole or in part from Transporter Classification Database. The copyright holder has licensed the content in a manner that permits reuse under CC BY-SA 3.0 and GFDL. All relevant terms must be followed. The original text was at "2.A.55 The Metal Ion (Mn2+-iron) Transporter (Nramp) Family"