
The webbed foot is a specialized limb with interdigital membranes (webbings) that aids in aquatic locomotion, present in a variety of tetrapod vertebrates. This adaptation is primarily found in semiaquatic species, and has convergently evolved many times across vertebrate taxa.

It likely arose from mutations in developmental genes that normally cause tissue between the digits to apoptose. These mutations were beneficial to many semiaquatic animals because the increased surface area from the webbing allowed for more swimming propulsion and swimming efficiency, especially in surface swimmers.[2] The webbed foot also has enabled other novel behaviors like escape responses and mating behaviors. A webbed foot may also be called a paddle to contrast it from a more hydrofoil-like flipper.
Morphology

A webbed foot has connecting tissue between the toes of the foot. Several distinct conditions can give rise to webbed feet, including interdigital webbing and syndactyly. The webbing can consist of membrane, skin, or other connective tissue and varies widely in different taxa. This modification significantly increases the surface area of the feet. One of the consequences of this modification in some species, specifically birds, is that the feet are a major location for heat loss.[3] In birds, the legs utilize countercurrent heat exchange so that blood reaching the feet is already cooled by blood returning to the heart to minimize this effect.[4][5] Webbed feet take on a variety of different shapes; in birds, the webbing can even be discontinuous, as seen in lobate-footed birds like grebes.[6] However, one of the most common is the delta (Δ) or triangular shape seen in most waterfowl and frogs.[1] This delta wing shape is a solution that has convergently evolved in many taxa, and is also used in aircraft to allow for high lift forces at high attack angles. This shape allows for the production of large forces during swimming through both drag-based and lift-based propulsion.[1]
Webbed feet are a compromise between aquatic and terrestrial locomotion. Aquatic control surfaces of non-piscine vertebrates may be paddles or hydrofoils. Paddles generate less lift than hydrofoils, and paddling is associated with drag-based control surfaces. The roughly triangular design of webbed feet, with a broad distal end, is specialized to increase propulsive efficiency by affecting a larger mass of water over generating increased lift. This is in contrast to a more hydrofoil-like flipper of many permanently aquatic animals.[7]
Evolution
Development
Webbed feet are the result of mutations in genes that normally cause interdigital tissue between the toes to apoptose.[8] Apoptosis, or programmed cell death, in development is mediated by a variety of pathways, and normally causes the creation of digits by death of tissue separating the digits. Different vertebrate species with webbed feet have different mutations that disrupt this process, indicating that the structure arose independently in these lineages.
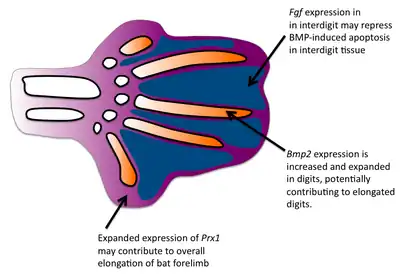
In humans, syndactyly can arise from as many as nine unique subtypes with their own clinical, morphological, and genetic fingerprints. In addition, the same genetic mutations can underlie different phenotypic expressions of syndactyly.[10] While these conditions are disorders in humans, the variability in genetic cause of webbed digits informs our understanding of how this morphological change arose in species where webbed feet were selectively advantageous. These conditions also demonstrate a variety of genetic targets for mutation resulting in webbed feet, which may explain how this homologous structure could have arisen many times over the course of evolutionary history.
One pathway implicated in interdigital necrosis is the bone morphogenetic protein (BMP) signaling pathway. BMP signaling molecules (BMPs) are expressed in the tissue regions between digits during development. In experiments with chickens, mutations to a BMP receptor disrupted the apoptosis of interdigital tissue and caused webbed feet similar to ducks to develop. In ducks, BMPs are not expressed at all.[11] These results indicate that in avian lineages, the disruption of BMP signaling in interdigital tissue caused webbed feet to arise. The magnitude of attenuation in this pathway is correlated with the amount of interdigital tissue preserved. Other genetic changes implicated in webbed feet development in avians include reduction of TGFβ-induced chondrogenesis and reduction of msx-1 and msx-2 gene expression.[12]
Webbed feet could also arise due to being linked to other morphological changes, without a selective advantage. In salamanders, webbed feet have arisen in multiple lineages, but in most do not contribute to increased function. However, in the cave salamander species Chiropterotriton magnipes (bigfoot splayfoot salamander), their webbed feet are morphologically unique from other salamanders and may serve a functional purpose.[13] This demonstrates that webbed feet arise from developmental changes, but do not necessarily correlate with a selective advantage functionally.
Phylogeny
Webbed feet have arisen in all major vertebrate lineages with limbed animals. Most webbed-footed species spend part of their time in aquatic environments, indicating that this homologous structure provides some advantage to swimmers. Some examples from each class are highlighted here, but this is not a complete listing.

Amphibians
Of the three orders of amphibians, Anura (frogs and toads) and Urodela (salamanders) have representative species with webbed feet. Frogs that live in aquatic environments, like the common frog (Rana temporaria), have webbed feet. Salamanders in arboreal and cave environments also have webbed feet, but in most species, this morphological change does not likely have a functional advantage.[13]
Reptiles
Reptiles have webbed-footed representatives that include freshwater turtles and geckos. While turtles with webbed feet are aquatic, most geckos live in terrestrial and arboreal environments.
Birds
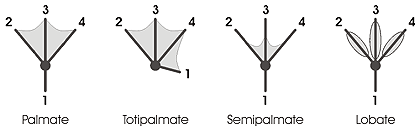
Birds are typically classified as a sub-group of reptiles, but they are a distinct class within vertebrates, so are discussed separately. Birds have a wide span of representatives with webbed feet, due to the diversity of waterfowl. Ducks, geese, and swans all have webbed feet. They utilize different foraging behaviors in water, but use similar modes of locomotion. There is a wide variety of webbing and lobation styles in bird feet, including birds with all digits joined in webbing, like the Brandt's cormorant and birds with lobed digits, like grebes. Palmations and lobes enable swimming or help walking on loose ground such as mud.[14] The webbed or palmated feet of birds can be categorized into several types:
- Palmate: only the anterior digits (2–4) are joined by webbing. Found in ducks, geese and swans, gulls and terns, and other aquatic birds (auks, flamingos, fulmars, jaegers, loons, petrels, shearwaters and skimmers).[15][16] Diving ducks also have a lobed hind toe (1), and gulls, terns and allies have a reduced hind toe.[17]
- Totipalmate: all four digits (1–4) are joined by webbing. Found in gannets and boobies, pelicans, cormorants, anhingas, frigatebirds, and tropicbirds. Some gannets have brightly colored feet used in display.[14][16]
- Semipalmate: a small web between the anterior digits (2–4). Found in some plovers (Eurasian dotterels) and sandpipers (semipalmated sandpipers, stilt sandpipers, upland sandpipers, greater yellowlegs and willet), avocet, herons (only two toes), all grouse, and some domesticated breeds of chicken. Plovers and lapwings have a vestigial hind toe (1), and sandpipers and their allies have a reduced and raised hind toe barely touching the ground. The sanderling is the only sandpiper having 3 toes (tridactyl foot).[14]
- Lobate: the anterior digits (2–4) are edged with lobes of skin. Lobes expand or contract when a bird swims. In grebes, coots, phalaropes, finfoots and some palmate-footed ducks on the hallux (1). Grebes have more webbing between the toes than coots and phalaropes.[15][18][16]
The palmate foot is most common.
Mammals

Some semiaquatic mammals have webbed feet. Most of these have interdigital webbing, as opposed to the syndactyly found in birds. Some notable examples include the platypus, the beaver, the otter, and the water opossum.[19][20][21]
Function
Swimming propulsion
In many species, webbed feet likely evolved to aid in generation of propulsion during swimming. Most webbed-footed animals utilize paddling modes of locomotion where their feet stroke backwards relative to their whole body motion, generating a propulsive force. The interdigital membrane increases the surface area, which increases the propulsive drag the animal can generate with each stroke of its foot.[22][23] This is a drag-based mode of propulsion. However, some waterfowl also utilize lift-based modes of propulsion, where their feet generate hydrodynamic lift due to the angle of attack of the foot and the relative water velocity. For example, great-crested grebes use solely lift-based propulsion due to their lateral foot stroke and asymmetric, lobated toes.[6] Most waterfowl use a combination of these two modes of propulsion, where the first third of their foot stroke generates propulsive drag and the last two-thirds of the stroke generates propulsive lift.[1]
The stroke of the foot through the water also generates vortices that aid propulsion. During the transition from drag-based to lift-based propulsion in ducks, leading edge vortices formed on the front of the foot are shed, which creates a flow of water over the foot that likely aids lift production.[1] Other species also create these vortices during their webbed foot stroke. Frogs also create vortices that shed off their feet when swimming in water. The vortices from the two feet do not interfere with each other; therefore, each foot is generating forward propulsion independently.[24]
Most fully aquatic vertebrates do not use paddling modes of locomotion, instead using undulatory modes of locomotion or flipper locomotion. Fully aquatic mammals and animals typically have flippers instead of webbed feet, which are a more heavily specialized and modified limb.[2] It is hypothesized that an evolutionary transition between semiaquatic and fully aquatic higher vertebrates (especially mammals) involved both the specialization of swimming limbs and the transition to underwater, undulatory modes of motion.[25] However, for semiaquatic animals that mainly swim at the surface, webbed feet are highly functional; they trade-off effectively between efficient terrestrial and aquatic locomotion.[2] In addition, some waterfowl can also use paddling modes for underwater swimming, with added propulsion from flapping their wings. Diving ducks can swim underwater to forage. These ducks expend more than 90% of their energy to overcome their own buoyancy when they dive.[26] They can also achieve higher speeds underwater due to surface speeds being limited to their hull speed; at this speed, the wave drag increases to the point where the duck cannot swim faster.[27]
Other behaviors
In ducks, webbed feet have also enabled extreme forms of propulsion that are used for escape behaviors and courtship display. Surface swimmers are speed-limited due to increasing drag as they approach a physically defined hull speed, which is determined by their body length. In order to achieve speeds higher than hull speed, some ducks, like eider ducks, use distinctive modes of locomotion that involve lifting the body out of the water. They can hydroplane, where they lift part of their body out of the water and paddle with their webbed feet to generate forces that allow them to overcome gravity; they also use paddle-assisted flying, where the whole body is lifted out of the water, and the wings and feet work in concert to generate lift forces.[28] In extreme cases, this type of behavior is used for sexual selection. Western and Clark's grebes utilize their lobated feet to generate nearly 50% of the force required to allow them to walk on water in elaborate sexual displays; they are likely the largest animal to "walk" on water, and are an order of magnitude heavier than the well-known lizards that exhibit a similar behavior.[29]
Terrestrial locomotion
While webbed feet have mainly arisen in swimming species, they can also aid in terrestrial locomotors by increasing contact area on slick or soft surfaces. For P. rangei, the Namib sand gecko, their webbed feet may serve as sand shoes that enable them to move atop sand dunes.[30] However, some ecologists believe that their webbed feet do not aid aboveground locomotion, but are mainly utilized as shovels for burrowing and digging in the sand.[31] In salamanders, most species do not benefit from the increased surface area of their feet. However, some, like the bigfoot splayfoot salamander (Chiropterotriton magnipes) increase their body size to foot surface area ratio enough to provide increased suction. This species lives in cave environments where they often encounter wet, slick surfaces. Therefore, their webbed feet may enable them to move on these surfaces with ease.[13]
See also
References
- 1 2 3 4 5 Johansson, L. Christoffer; Norberg, R. Ake (2003-07-03). "Delta-wing function of webbed feet gives hydrodynamic lift for swimming propulsion in birds". Nature. 424 (6944): 65–68. Bibcode:2003Natur.424...65J. doi:10.1038/nature01695. ISSN 1476-4687. PMID 12840759. S2CID 4429458.
- 1 2 3 Fish, F. E. (1984-05-01). "Mechanics, power output and efficiency of the swimming muskrat (Ondatra zibethicus)". The Journal of Experimental Biology. 110: 183–201. doi:10.1242/jeb.110.1.183. ISSN 0022-0949. PMID 6379093.
- ↑ "Webbed Wonders". www.ducks.org. Retrieved 2017-04-17.
- ↑ Gill, Frank B. (1994). Ornithology. ISBN 978-0716724155. OCLC 959809850.
- ↑ "Why Don't Ducks' Feet Freeze?". Ask a Naturalist.com. 2010-04-22. Retrieved 2017-04-18.
- 1 2 Johansson, L. C.; Norberg, U. M. (2000-10-05). "Asymmetric toes aid underwater swimming". Nature. 407 (6804): 582–583. doi:10.1038/35036689. ISSN 0028-0836. PMID 11034197. S2CID 4302176.
- ↑ Fish, F.E. (2004). "Structure and Mechanics of Nonpiscine Control Surfaces". IEEE Journal of Oceanic Engineering. 29 (3): 605–621. Bibcode:2004IJOE...29..605F. doi:10.1109/joe.2004.833213. ISSN 0364-9059. S2CID 28802495.
- ↑ Sadava, David E.; Orians, Gordon H.; Heller, H. Craig; Hillis, David M.; Purves, William K. (2006-11-15). Life (Loose Leaf): The Science of Biology. Macmillan. ISBN 9781429204590.
- ↑ Weatherbee, Scott D.; Behringer, Richard R.; Rasweiler, John J.; Niswander, Lee A. (2006-10-10). "Interdigital webbing retention in bat wings illustrates genetic changes underlying amniote limb diversification". Proceedings of the National Academy of Sciences of the United States of America. 103 (41): 15103–15107. Bibcode:2006PNAS..10315103W. doi:10.1073/pnas.0604934103. ISSN 0027-8424. PMC 1622783. PMID 17015842.
- ↑ Malik, Sajid (2017-04-27). "Syndactyly: phenotypes, genetics and current classification". European Journal of Human Genetics. 20 (8): 817–824. doi:10.1038/ejhg.2012.14. ISSN 1018-4813. PMC 3400728. PMID 22333904.
- ↑ Zou, Hongyan; Niswander, Lee (1996-01-01). "Requirement for BMP Signaling in Interdigital Apoptosis and Scale Formation". Science. 272 (5262): 738–741. Bibcode:1996Sci...272..738Z. doi:10.1126/science.272.5262.738. JSTOR 2889452. PMID 8614838. S2CID 27174863.
- ↑ Gañan, Yolanda; Macias, Domingo; Basco, Ricardo D.; Merino, Ramón; Hurle, Juan M. (1998). "Morphological Diversity of the Avian Foot Is Related with the Pattern of msx Gene Expression in the Developing Autopod". Developmental Biology. 196 (1): 33–41. doi:10.1006/dbio.1997.8843. PMID 9527879.
- 1 2 3 Jaekel, Martin; Wake, David B. (2007-12-18). "Developmental processes underlying the evolution of a derived foot morphology in salamanders". Proceedings of the National Academy of Sciences of the United States of America. 104 (51): 20437–20442. Bibcode:2007PNAS..10420437J. doi:10.1073/pnas.0710216105. ISSN 0027-8424. PMC 2154449. PMID 18077320.
- 1 2 3 Kochan 1994; Proctor & Lynch 1993; Elphick, Dunning & Sibley 2001
- 1 2 Gill 2001; Kochan 1994; Proctor & Lynch 1993; Elphick, Dunning & Sibley 2001
- 1 2 3 Kalbe, Lothar (1983). "Besondere Formen für spezielle Aufgaben der Wassertiere [Special adaptations of aquatic animals to specific lifestyles]". Tierwelt am Wasser [Wildlife by the Water] (in German) (1st ed.). Leipzig-Jena-Berlin: Urania-Verlag. pp. 72–77.
- ↑ Kochan 1994; Elphick, Dunning & Sibley 2001
- ↑ Kowalska-Dyrcz, Alina (1990). "Entry: noga [leg]". In Busse, Przemysław (ed.). Ptaki [Birds]. Mały słownik zoologiczny [Small zoological dictionary] (in Polish). Vol. I (I ed.). Warsaw: Wiedza Powszechna. pp. 383–385. ISBN 978-83-214-0563-6.
- ↑ Fish, F. E.; Baudinette, R. V.; Frappell, P. B.; Sarre, M. P. (1997). "Energetics of Swimming by the Platypus Ornithorhynchus anatinus: Metabolic Effort Associated with Rowing" (PDF). The Journal of Experimental Biology. 200 (20): 2647–52. doi:10.1242/jeb.200.20.2647. PMID 9359371.
- ↑ Yadav, P. R.; Khanna, D. R. (2005). Biology of Mammals. Discovery Publishing House. p. 124. ISBN 978-8171419340.
- ↑ Muller-Schwarze, Dietland; Sun, Lixing (2003). The Beaver: Natural History of a Wetlands Engineer. Comstock Publishing Associates. p. 12. ISBN 978-0801440984.
- ↑ Thewissen, J. G. M. (1998-10-31). The Emergence of Whales: Evolutionary Patterns in the Origin of Cetacea. Springer Science & Business Media. ISBN 9780306458538.
- ↑ Lulashnyk, Lorne (2016-12-19). Understanding Surfaces. FriesenPress. ISBN 9781460274309.
- ↑ Stamhuis, Eize J.; Nauwelaerts, Sandra (2005-04-01). "Propulsive force calculations in swimming frogs. II. Application of a vortex ring model to DPIV data" (PDF). The Journal of Experimental Biology. 208 (Pt 8): 1445–1451. doi:10.1242/jeb.01530. ISSN 0022-0949. PMID 15802668. S2CID 34494254.
- ↑ Fish, Frank E. (1994-01-01). "Association of Propulsive Swimming Mode with Behavior in River Otters (Lutra canadensis)". Journal of Mammalogy. 75 (4): 989–997. doi:10.2307/1382481. JSTOR 1382481.
- ↑ Ribak, Gal; Swallow, John G.; Jones, David R. (2010-09-07). "Drag-based 'hovering' in ducks: the hydrodynamics and energetic cost of bottom feeding". PLOS ONE. 5 (9): e12565. Bibcode:2010PLoSO...512565R. doi:10.1371/journal.pone.0012565. ISSN 1932-6203. PMC 2935360. PMID 20830286.
- ↑ Ancel, A.; Starke, L. N.; Ponganis, P. J.; Van Dam, R.; Kooyman, G. L. (2000-12-01). "Energetics of surface swimming in Brandt's cormorants (Phalacrocorax penicillatus Brandt)". The Journal of Experimental Biology. 203 (Pt 24): 3727–3731. doi:10.1242/jeb.203.24.3727. ISSN 0022-0949. PMID 11076736.
- ↑ Gough, William T.; Farina, Stacy C.; Fish, Frank E. (2015-06-01). "Aquatic burst locomotion by hydroplaning and paddling in common eiders (Somateria mollissima)". The Journal of Experimental Biology. 218 (Pt 11): 1632–1638. doi:10.1242/jeb.114140. ISSN 1477-9145. PMID 25852065.
- ↑ Clifton, Glenna T.; Hedrick, Tyson L.; Biewener, Andrew A. (2015-04-15). "Western and Clark's grebes use novel strategies for running on water". The Journal of Experimental Biology. 218 (Pt 8): 1235–1243. doi:10.1242/jeb.118745. ISSN 1477-9145. PMID 25911734.
- ↑ "Web-Footed Geckos". National Geographic. Archived from the original on February 7, 2010. Retrieved 2017-04-28.
- ↑ Russell, A. P.; Bauer, A. M. (1990-12-01). "Substrate excavation in the Namibian web-footed gecko, Palmatogecko rangei Andersson 1908, and its ecological significance". Tropical Zoology. 3 (2): 197–207. doi:10.1080/03946975.1990.10539462. ISSN 0394-6975.
Sources
- Elphick, John B.; Dunning, Jack B. Jr.; Sibley, David Allen (2001). National Audubon Society: The Sibley Guide to Bird Life & Behavior. New York: Alfred A. Knopf. ISBN 978-0-679-45123-5.
- Gill, Frank B. (2001). Ornithology (2nd ed.). New York: W.H. Freeman and Company. ISBN 978-0-7167-2415-5.
- Kochan, Jack B. (1994). Feet & Legs. Birds. Mechanicsburg: Stackpole Books. ISBN 978-0-8117-2515-6.
- Proctor, Noble S.; Lynch, Patrick J. (1993). "Chapters: 6. Topography of the foot, 11. The pelvic girdle, and 12. The bones of the leg and foot Family". Manual of Ornithology. Avian Structure & Function. New Haven and London: Yale University Press. pp. 70–75, 140–141, 142–144. ISBN 978-0-300-07619-6.
