| Presenilin | |||||||||
|---|---|---|---|---|---|---|---|---|---|
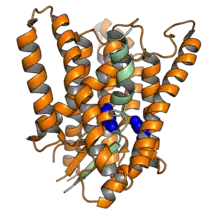 Cryo-electron microscopy structure of the human presenilin-1 (orange) in complex with a fragment of one of its protein substrates, Notch (green). The two catalytic sites are shown in blue. Rendered from PDB: 6IDF.[1] | |||||||||
| Identifiers | |||||||||
| Symbol | Presenilin | ||||||||
| Pfam | PF01080 | ||||||||
| Pfam clan | CL0130 | ||||||||
| InterPro | IPR001108 | ||||||||
| MEROPS | A22 | ||||||||
| TCDB | 1.A.54 | ||||||||
| OPM superfamily | 244 | ||||||||
| OPM protein | 4hyg | ||||||||
| |||||||||
| presenilin 1 (Alzheimer's disease 3) | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Symbol | PSEN1 | ||||||
| Alt. symbols | AD3 | ||||||
| NCBI gene | 5663 | ||||||
| HGNC | 9508 | ||||||
| OMIM | 104311 | ||||||
| RefSeq | NM_000021 | ||||||
| UniProt | P49768 | ||||||
| Other data | |||||||
| EC number | 3.4.23.- | ||||||
| Locus | Chr. 14 q24.3 | ||||||
| |||||||
| presenilin 2 (Alzheimer's disease 4) | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Symbol | PSEN2 | ||||||
| Alt. symbols | AD4 | ||||||
| NCBI gene | 5664 | ||||||
| HGNC | 9509 | ||||||
| OMIM | 600759 | ||||||
| RefSeq | NM_000447 | ||||||
| UniProt | P49810 | ||||||
| Other data | |||||||
| EC number | 3.4.23.- | ||||||
| Locus | Chr. 1 q31-q42 | ||||||
| |||||||
Presenilins are a family of related multi-pass transmembrane proteins which constitute the catalytic subunits of the gamma-secretase intramembrane protease protein complex. They were first identified in screens for mutations causing early onset forms of familial Alzheimer's disease by Peter St George-Hyslop.[2] Vertebrates have two presenilin genes, called PSEN1 (located on chromosome 14 in humans) that codes for presenilin 1 (PS-1) and PSEN2 (on chromosome 1 in humans) that codes for presenilin 2 (PS-2).[3] Both genes show conservation between species, with little difference between rat and human presenilins. The nematode worm C. elegans has two genes that resemble the presenilins and appear to be functionally similar, sel-12 and hop-1.[4]
Presenilins undergo cleavage in an alpha helical region of one of the cytoplasmic loops to produce a large N-terminal and a smaller C-terminal fragment that together form part of the functional protein.[5] Cleavage of presenilin 1 can be prevented by a mutation that causes the loss of exon 9, and results in loss of function. Presenilins play a key role in the modulation of intracellular Ca2+ involved in presynaptic neurotransmitter release and long-term potentiation induction.[6]
Structure
Presenilins are transmembrane proteins with nine alpha helices. Structures have been solved of the assembled gamma secretase complex by cryo-electron microscopy, demonstrating significant conformational flexibility in the structure of the presenilin subunit of the complex in response to ligand or inhibitor binding.[7][1] Presenilins undergo autocatalytic proteolytic processing after expression, cleaving a cytoplasmic loop region between the sixth and seventh helices to produce a large N-terminal and a smaller C-terminal fragment. The two fragments remain in contact with each other in the mature protein. The two catalytic aspartate active site residues required for aspartyl protease activity are located in the sixth and seventh helices.[8]
The structure and membrane topology of presenilins was originally controversial when they were first discovered. The PSEN1 gene was predicted to contain ten trans-membrane domains; models agreed on the expected topology of the N-terminal fragment, but the structure of the C-terminal fragment was disputed. A 2006 study suggested a nine-pass transmembrane topology with cleavage and assembly into the gamma-secretase complex prior to insertion into the plasma membrane.[9] Solution NMR studies of the C-terminal fragment showed three helices likely to traverse the membrane,[5][10] while X-ray crystallography studies of an archaeal homolog,[11] as well as cryo-electron microscopy of the human gamma-secretase complex, indicate nine transmembrane helices.[7][1]
Function
Catalytic
Presenilins are the catalytic component of the gamma secretase intramembrane protease, a four-member protein complex consisting of presenilin, nicastrin, APH-1, and PEN-2. It has a very broad range of substrates for its proteolytic activity. Its substrates are hydrophobic single-pass transmembrane helices with relatively small extracellular regions.[12][13] These substrates arise following ectodomain shedding.[13] Well over 100 different integral membrane proteins are processed by gamma-secretase.[8] The best-characterized gamma-secretase substrates are the Notch receptor and amyloid precursor protein (APP).[7][13] Presenilins' role in the Notch signaling pathway is important in development;[14] mice that have the PS1 gene knocked out die early in development from developmental abnormalities similar to those found when notch is disrupted.[15] In conditional knockout mice where presenilin is only inactivated after early development, there is evidence that presenilins in their role as gamma-secretase components are important in the survival of neurons during aging.[14]
There are subtle and species-specific variations in the roles of presenilin-1 and presenilin-2 in assembled gamma-secretase complexes, with many studies suggesting a primary role for presenilin-1.[13] In humans, the two presenilins differ in subcellular localization, and may also be cell type and tissue-specific.[8]
Non-catalytic
Presenilins also have additional non-catalytic roles in other cellular signaling processes, including calcium homeostasis, lysosomal acidification, autophagy, and protein trafficking.[16][17][18] The proteins' role in calcium homeostasis in neurons has been a subject of interest.[19] The genetic inactivation of presenilins in hippocampal synapses has shown this selectively affects the long-term potentiation caused by theta with the inactivation in presynapse but not the postsynapse impairing short-term plasticity and synaptic facilitation.[6] The release of glutamate was also reduced in presynaptic terminals by processes that involve modulation of intracellular Ca2+ release.[6] This has been suggested to "represent a general convergent mechanism leading to neurodegeneration".[6]
Homologs have been identified and characterized in diverse eukaryotic organisms, including model organisms Drosophila melanogaster and Caenorhabditis elegans, plants such as Arabidopsis thaliana and Physcomitrella patens, and the slime mold Dictyostelium discoideum.[17][18] In these functions presenilins are thought to play a role as scaffold proteins, considered likely to be the ancestral role of the protein family.[18]
Expression and distribution
Both human presenilins have widespread expression in the brain. The two proteins differ in subcellular localization, with PS1 expressed more broadly and present at the cell membrane, while PS2 is present mainly in late endosomes and lysosomes.[8][20]
Association with Alzheimer's disease
Most cases of Alzheimer's disease are not hereditary. However, there is a small subset of cases that have an earlier age of onset and have a strong genetic element. In patients with Alzheimer's disease (autosomal dominant hereditary), mutations in the presenilin proteins (PSEN1; PSEN2) or the amyloid precursor protein (APP) can be found. The majority of these cases carry mutant presenilin genes. An important part of the disease process in Alzheimer's disease is the accumulation of Amyloid beta (Aβ) protein. To form Aβ, APP must be cut by two enzymes, beta secretases and gamma secretase. Presenilin is the sub-component of gamma secretase that is responsible for the cutting of APP.
Gamma secretase can cut APP at several points within a small region of the protein, which results in Aβ of various lengths. The lengths associated with Alzheimer's disease are 40 and 42 amino acids long. Aβ 42 is more likely to aggregate to form plaques in the brain than Aβ 40. Presenilin mutations lead to an increase in the ratio of Aβ 42 produced compared to Aβ 40, although the total quantity of Aβ produced remains constant.[21] This can come about by various effects of the mutations upon gamma secretase.[22]
Discovery
The genes for the presenilins were discovered in 1995 through linkage studies using mutations present in familial Alzheimer's disease cases.[2] Around the same time, the presenilin homolog in Caenorhabditis elegans, sel-12, was independently identified as a contributor to Notch signaling.[23] Although the function of the protein products of these genes was not immediately apparent, it became clear from subsequent work that the mutations were associated with higher proportions of Aβ42 over the less amyloidogenic Aβ40. The role of presenilins as the catalytic component of the gamma secretase protein complex was established by the early 2000s.[24][25]
References
- 1 2 3 Yang G, Zhou R, Zhou Q, Guo X, Yan C, Ke M, et al. (January 2019). "Structural basis of Notch recognition by human γ-secretase". Nature. 565 (7738): 192–197. doi:10.1038/s41586-018-0813-8. PMID 30598546. S2CID 57189683.
- 1 2 Sherrington R, Rogaev EI, Liang Y, Rogaeva EA, Levesque G, Ikeda M, et al. (June 1995). "Cloning of a gene bearing missense mutations in early-onset familial Alzheimer's disease". Nature. 375 (6534): 754–60. Bibcode:1995Natur.375..754S. doi:10.1038/375754a0. PMID 7596406. S2CID 4308372.
- ↑ Levy-Lahad E, Wasco W, Poorkaj P, Romano DM, Oshima J, Pettingell WH, et al. (August 1995). "Candidate gene for the chromosome 1 familial Alzheimer's disease locus". Science. 269 (5226): 973–7. Bibcode:1995Sci...269..973L. doi:10.1126/science.7638622. PMID 7638622. S2CID 27296868.
- ↑ Smialowska A, Baumeister R (2006). "Presenilin function in Caenorhabditis elegans". Neuro-Degenerative Diseases. 3 (4–5): 227–32. doi:10.1159/000095260. PMID 17047361. S2CID 9695127.
- 1 2 Sobhanifar S, Schneider B, Löhr F, Gottstein D, Ikeya T, Mlynarczyk K, et al. (May 2010). "Structural investigation of the C-terminal catalytic fragment of presenilin 1". Proceedings of the National Academy of Sciences of the United States of America. 107 (21): 9644–9. Bibcode:2010PNAS..107.9644S. doi:10.1073/pnas.1000778107. PMC 2906861. PMID 20445084.
- 1 2 3 4 Zhang C, Wu B, Beglopoulos V, Wines-Samuelson M, Zhang D, Dragatsis I, et al. (July 2009). "Presenilins are essential for regulating neurotransmitter release". Nature. 460 (7255): 632–6. Bibcode:2009Natur.460..632Z. doi:10.1038/nature08177. PMC 2744588. PMID 19641596.
- 1 2 3 Bai XC, Rajendra E, Yang G, Shi Y, Scheres SH (December 2015). "Sampling the conformational space of the catalytic subunit of human γ-secretase". eLife. 4: e11182. doi:10.7554/eLife.11182. PMC 4718806. PMID 26623517.
- 1 2 3 4 Güner G, Lichtenthaler SF (September 2020). "The substrate repertoire of γ-secretase/presenilin". Seminars in Cell & Developmental Biology. 105: 27–42. doi:10.1016/j.semcdb.2020.05.019. PMID 32616437.
- ↑ Spasic D, Tolia A, Dillen K, Baert V, De Strooper B, Vrijens S, Annaert W (September 2006). "Presenilin-1 maintains a nine-transmembrane topology throughout the secretory pathway". The Journal of Biological Chemistry. 281 (36): 26569–77. doi:10.1074/jbc.M600592200. PMID 16846981.
- ↑ PDB: 2KR6; Doetsch V (2010). "Solution structure of presenilin-1 CTF subunit". Worldwide Protein Data Bank. doi:10.2210/pdb2kr6/pdb.
- ↑ Li X, Dang S, Yan C, Gong X, Wang J, Shi Y (January 2013). "Structure of a presenilin family intramembrane aspartate protease". Nature. 493 (7430): 56–61. Bibcode:2013Natur.493...56L. doi:10.1038/nature11801. PMID 23254940. S2CID 4429356.
- ↑ Struhl G, Adachi A (September 2000). "Requirements for presenilin-dependent cleavage of notch and other transmembrane proteins". Molecular Cell. 6 (3): 625–36. doi:10.1016/S1097-2765(00)00061-7. PMID 11030342.
- 1 2 3 4 Stanga S, Vrancx C, Tasiaux B, Marinangeli C, Karlström H, Kienlen-Campard P (February 2018). "Specificity of presenilin-1- and presenilin-2-dependent γ-secretases towards substrate processing". Journal of Cellular and Molecular Medicine. 22 (2): 823–833. doi:10.1111/jcmm.13364. PMC 5783875. PMID 28994238.
- 1 2 Shen J (2 October 2013). "Function and dysfunction of presenilin". Neuro-Degenerative Diseases. 13 (2–3): 61–3. doi:10.1159/000354971. PMC 4000081. PMID 24107444.
- ↑ Shen J, Bronson RT, Chen DF, Xia W, Selkoe DJ, Tonegawa S (May 1997). "Skeletal and CNS defects in Presenilin-1-deficient mice". Cell. 89 (4): 629–39. doi:10.1016/S0092-8674(00)80244-5. PMID 9160754. S2CID 8252011.
- ↑ Peric A, Annaert W (March 2015). "Early etiology of Alzheimer's disease: tipping the balance toward autophagy or endosomal dysfunction?". Acta Neuropathologica. 129 (3): 363–81. doi:10.1007/s00401-014-1379-7. PMC 4331606. PMID 25556159.
- 1 2 Duggan SP, McCarthy JV (January 2016). "Beyond γ-secretase activity: The multifunctional nature of presenilins in cell signalling pathways". Cellular Signalling. 28 (1): 1–11. doi:10.1016/j.cellsig.2015.10.006. PMID 26498858.
- 1 2 3 Otto GP, Sharma D, Williams RS (April 2016). "Non-Catalytic Roles of Presenilin Throughout Evolution". Journal of Alzheimer's Disease. 52 (4): 1177–87. doi:10.3233/JAD-150940. PMC 4927835. PMID 27079701.
- ↑ Honarnejad K, Herms J (November 2012). "Presenilins: role in calcium homeostasis". The International Journal of Biochemistry & Cell Biology. 44 (11): 1983–6. doi:10.1016/j.biocel.2012.07.019. PMID 22842534.
- ↑ Barthet G, Mulle C (November 2020). "Presynaptic failure in Alzheimer's disease" (PDF). Progress in Neurobiology. 194: 101801. doi:10.1016/j.pneurobio.2020.101801. PMID 32428558. S2CID 218655934.
- ↑ Citron M, Westaway D, Xia W, Carlson G, Diehl T, Levesque G, et al. (January 1997). "Mutant presenilins of Alzheimer's disease increase production of 42-residue amyloid beta-protein in both transfected cells and transgenic mice". Nature Medicine. 3 (1): 67–72. doi:10.1038/nm0197-67. PMID 8986743. S2CID 10834438.
- ↑ Bentahir M, Nyabi O, Verhamme J, Tolia A, Horré K, Wiltfang J, et al. (February 2006). "Presenilin clinical mutations can affect gamma-secretase activity by different mechanisms". Journal of Neurochemistry. 96 (3): 732–42. doi:10.1111/j.1471-4159.2005.03578.x. PMID 16405513.
- ↑ Levitan D, Greenwald I (September 1995). "Facilitation of lin-12-mediated signalling by sel-12, a Caenorhabditis elegans S182 Alzheimer's disease gene". Nature. 377 (6547): 351–4. Bibcode:1995Natur.377..351L. doi:10.1038/377351a0. PMID 7566091. S2CID 4314257.
- ↑ De Strooper B, Iwatsubo T, Wolfe MS (January 2012). "Presenilins and γ-secretase: structure, function, and role in Alzheimer Disease". Cold Spring Harbor Perspectives in Medicine. 2 (1): a006304. doi:10.1101/cshperspect.a006304. PMC 3253024. PMID 22315713.
- ↑ Wolfe MS (February 2007). "When loss is gain: reduced presenilin proteolytic function leads to increased Abeta42/Abeta40. Talking Point on the role of presenilin mutations in Alzheimer disease". EMBO Reports. 8 (2): 136–40. doi:10.1038/sj.embor.7400896. PMC 1796780. PMID 17268504.
External links
- The MEROPS online database for peptidases and their inhibitors: Presenilin 1 A22.001, Presenilin 2 A22.002
- Dominantly Inherited Alzheimer Network (DIAN)
- Dominantly Inherited Alzheimer Network (DIAN) Expanded Registry
- Presenilins at the U.S. National Library of Medicine Medical Subject Headings (MeSH)