四足類
四足類(英語:)是一支主要为适应陆地环境而演化的有颌类脊椎动物,在生物分类学上组成了四足总纲或四足超纲(學名:),包含两栖动物和羊膜动物两支,后者包括合弓纲(哺乳类及其已灭绝的史前亲族)和蜥形纲(爬行类、鸟类和其已灭绝的史前亲族)两大演化支。四足动物是四足形類的冠群,由泥盆紀晚期的肉鳍鱼类演化而来,是肉鳍鱼现存的主要后裔,包括除少数两栖鱼类之外的所有陆生和半水生脊椎动物。
| 四足總綱 | |
|---|---|
 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 高纲: | 硬骨鱼高纲 Osteichthyes |
| 演化支: | 肉鳍鱼类 Sarcopterygii |
| 演化支: | 四足形類 Tetrapodomorpha |
| 演化支: | 始四足类 Eotetrapodiformes |
| 演化支: | 希望螈类 Elpistostegalia |
| 演化支: | 堅頭類 Stegocephalia |
| 总纲: | 四足總綱 Tetrapoda Jaekel, 1909[2] |
| 現存分支 | |
| 古生物 |
| 古生物学 |
|---|
 |
|
Portal:生物学 Category:古生物学 |
作为陆生演化支,四足类动物的成体都用与鱼鳔同源的肺直接呼吸空气进行气体交换,其中两栖类在幼体阶段为水生会暂时使用鳃呼吸,有些物种(两栖类和少数爬行类)在成年后还可以通过皮肤或肠道呼吸。如其名所示,大部分四足类都擁有左右对称的前后两对共四个肢体(包括鳍肢和翅膀等特化肢体),同源对应鱼类的胸鳍和腹鳍,且每个肢体都由有关节相连的内骨骼支撑并配带骨骼肌。但也有一些物种(蛇类、蛇蜥和蚓螈)因为Hox基因变异在演化过程中退化失去了四肢,仅在少数群体中残留肉眼可见的痕迹器官[3]。
多样性
| IUCN 对现存四足动物有关数据的估计(2014年)[4] | ||||||
|---|---|---|---|---|---|---|
| 种类 | 图片 | 纲 | 估计已描述的 物种数量[4] |
在 IUCN 红皮书中的 受威胁物种数量[4] |
已描述物种 百分比[4] |
受威胁物种 百分比[4] |
| 非羊膜动物 (在水中产卵) |
 |
两栖纲 (滑体类) |
7302 | 1957 | 88% | 41% |
| 羊膜动物 (在陆地产卵) |
 |
蜥形纲 (爬行类+鸟类) |
20463 | 2300 | 75% | 13% |
.png.webp) |
合弓纲 (哺乳类) |
5513 | 1199 | 100% | 26% | |
| 合计 | 33278 | 5456 | 80% | ? | ||
演化歷程
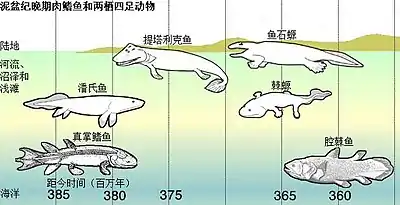
泥盆紀
詹妮弗·克拉克(Jennifer A. Clack)的研究顯示早期的四足類,如棘石螈(棘螈),是完全水中生活的,且並不適合陸地生活。這推翻了早期認為魚類因尋找獵物或乾涸首先進入陸地,再而演化出腳的說法。
第一個四足類相信是於泥盆紀晚期,即3.65亿年前(365百万年前)在淺水及沼澤環境下演化的。在泥盆紀晚期,陸地植物穩定了淡水環境,讓世界上首個濕地生態系統發展,加上不斷複雜化的食物網產生了更多的機會。[5]淡水環境並非唯一充滿有機物及植物的地方,像淺水濕地、海岸礁湖及大型的河流三角洲等的沼澤環境都十分普遍,這些都是適合四足類演化的地方。早期四足類化石都是在水中沉積岩中發現,加上一般原始四足類都分散在世界各地,所以牠們應該是沿海岸線分散,而並非只生活在淡水中。
肉鰭魚類演化出原始的四足類,牠那扁平的頭顱骨中有兩葉腦部、闊口及短鼻。牠們那雙向上看的眼睛可見牠們生活在河底。牠們的鰭發展出具適應性的肉鰭及骨骼。
潘氏魚是四足類的近親,有著鼻後孔。牠們生活在佈滿植物及碎石的淺水地區,以鰭為槳。牠們亦會用鰭來抓住植物,等待伏擊獵物。四足類的共同特徵是前肢在肘位向後彎曲,而後肢在膝蓋位置向前彎曲,在早期生活在淺水區的四足類都可以見到這些特徵。
硬骨魚的共同祖先有著原始的肺,可見牠們是從暖淺水中演化。這個環境正是肉鰭魚類生活的地方,並會當氧含量較低時利用牠們簡單構造的肺來呼吸。肺魚類現被認為是四足類的最近親。
肉鰭及骨頭是硬骨魚的原有特徵。四足類的肉鰭祖先進一步的演化,而輻鰭魚的祖先則從相反方向演化。最原始輻鰭魚的多鰭魚目就仍有肉質的前鰭。
泥盆紀共有9屬的基干四足類,有幾類主要或完全是基於下顎遺骸來描述。大部分都是在歐洲、格陵蘭及北美洲的超級大陸發現。另外在澳洲亦有發現,屬於岡瓦那大陸的澳洲螈。首種在亞洲發現的泥盆紀四足類是中國的潘氏中國螈。





最早的四足類並非陸上動物,而最早已知的陸地形態要追溯至石炭紀。牠們可能只會離開水面一段短時間,利用四肢來撥開汙泥。牠們為何離開水面仍被受爭議。其一可能是其幼體完成了適合陸地生活的變態:呼吸空氣及留在淺水近岸處作為保護,另外幼體正在兩個不同生態位的重疊處。其中一個生態位可能過於擠逼及危險,而另一個則較安全及不怎麼擠逼,對資源較小競爭。陸上生態位對於水中動物是具挑戰性的,但由於演化及物競天擇的壓力,能適應的幼體就能成功生存。一旦牠們能往陸地爬上一步,演化就會開始運作。
於此時在陸地上近水邊及潮濕的泥土中有很多無脊椎動物,足以成為四足類的食物。有些無脊椎動物甚至可以吃細小的四足類,但始終陸土是較為安全,且比水中能提供更多的資源。
成體要進入陸地實在太重及需要更大的獵物。細小的幼體則較輕、靈活及滿足於細小的無脊椎動物。現今的彈塗魚縱然在陸地上仍能夠捕捉飛行中的昆蟲,故此早期四足類幼體的生存能力不容忽視。
最初牠們只是臨時進入陸地,隨著一代一代的適應陸地環境及可以離開水面更長的時間,牠們的幼體開始更多時間在陸地生活。早期四足類成體亦有可能像魚石螈般因骨骼改變而開始進入陸地,但只是在岸邊曬太陽。事實上,第一種真正四足類能在陸地運動的都很細小,隨後才逐漸變大。
完全成長的四足類保有幼體的身體特徵及對陸地的適應性,並繼續改變四肢。一些細小物種的成體有可能可以在陸地上行走。
石炭紀
在1990年代前,泥盆紀晚期四足類與石炭紀中期兩棲類分支之間的化石紀錄有3000萬年的空白期,稱為「柔默空缺」。在這個空白期中,四足類發展了脊骨,四肢有趾及其他陸地上的適應性。耳朵、頭顱骨及脊柱都出現了變化。手指及腳趾的數目統一化為5隻,較多趾數的分支則滅絕。在空白期發現的僅有化石非常寶貴。
由水中的肉鰭魚類轉化為呼吸空氣的兩棲類在脊椎動物的演化歷史中是一重大事件。對於生活在中性重力及水中的環境的動物要進入陸地,對於身體的形態及功能要有完全的改變。引螈是一個例子。牠保存及重整了很多魚類祖先的特徵,強健的四肢可以支撐及運送離開水面的身體。較厚及強壯的脊骨防止身體因重量而下垂。再者,利用退化的魚類顎骨,發展了初步的耳朵,讓引螈可以聽到空氣傳送的聲音。
在石炭紀中期,早期四足類已演化成最少三個主要分支。離片椎類及爬行形类是四足類基底的代表。現今兩棲類是由離片椎類演化而來。首種羊膜動物是在石炭紀晚期出現。現存的四足類代表了這兩個分支的系統發生終點。第三個較原始的是勞克螈科,但卻已經滅絕。殼椎亞綱則是古生代的爬行形类。
分類
正真口類/硬骨鱼高纲 |
| ||||||||||||||||||||||||
| Osteichthyes |
四足類在四足形類中的系统发生位置(參考自2012年加州大學學者 Swartz 發表的論文[7]):
| 四足形類 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Tetrapodomorpha |
所有早期四足類及四足形類並非真正的兩棲類或羊膜動物,且曾被分類為迷齒亞綱的側系群。迷齒亞綱的特徵是牠們那複雜及象牙質包裹的牙齒結構,在肉鰭魚類中亦有同樣的特徵。迷齒亞綱可以分為魚石螈目、離片椎類及石炭蜥目。這三類的分別在於其脊骨結構。石炭蜥目有細小及融合的側椎體,在後期的脊椎動物成為脊柱。相反,離片椎類的側椎體則維持細小及原始的形態,並逐漸完全消失,椎間體很大及是完整的環。

四足類的部分分類如下:
- 脊椎动物亚门 Vertebrata
- 肉鳍鱼总纲 Sarcopterygii
- 四足形類 Tetrapodomorpha
- †散步鱼科 Elginerpetontidae
- †散步鱼 Elginerpeton pancheni Ahlberg, 1995(泥盆纪)
- †奥氏螈 Obruchevichthys gracilis Vorobyeva, 1977(泥盆纪)
- †厚颌螈 Densignathus rowei Daeschler, 2000(泥盆纪)
- †变额螈 Metaxygnathus denticulus Campbell & Bell, 1977(泥盆纪)
- †孔螈 Ventastega curonica Ahlberg, Luksevics & Lebedev, 1994(泥盆纪)
- †中国螈 Sinostega pani Zhu et al., 2002(泥盆纪)
- †古氏棘螈 Acanthostega gunnari Jarvik, 1952(泥盆纪)
- †魚石螈属 Ichthyostega Säve-Söderbergh, 1932(泥盆纪)
- †I. stensioi Säve-Söderbergh, 1932
- †I. eigili Säve-Söderbergh, 1932
- †I. watsoni Säve-Söderbergh, 1932
- †I. kochi Säve-Söderbergh, 1932
- †海納螈 Hynerpeton bassetti Daeschler et al., 1994(泥盆纪)
- †雅各布森螈 Jakubsonia livnensis Lebedev, 2004(泥盆纪)
- †于默螈 Ymeria denticulata Clack et al., 2012(泥盆纪)
- †图拉螈科 Tulerpetontidae
- †图拉螈 Tulerpeton. curtum Lebedeu, 1984(泥盆纪)
- †瓦切螈科 Whatcheeriidae
- †圆螈科 Colosteidae
- †厚蛙螈科 Crassigyrinidae
- †厚蛙螈 Crassigyrinus scoticus
- †斜眼螈总科 Baphetoidea
- †斜眼螈科 Baphetidae
- †斜眼螈亚科 Baphetinae
- †勞克螈亚科 Loxommatinae
- †斜眼螈科 Baphetidae
- 四足类 Tetrapoda
- †西格妮螈 Sigournea multidentata Bolt & Lombard, 2006
- †西方螈 Occidens portlocki Clack & Ahlberg, 2004
- 蛙形类 Batrachomorpha
- 爬行形類 Reptiliomorpha
- †散步鱼科 Elginerpetontidae
- 四足形類 Tetrapodomorpha
- 肉鳍鱼总纲 Sarcopterygii
早期四足類的身體特徵
四足類的魚類祖先必須擁有早期四足類的特徵,包括內鼻孔及有骨的大肉鰭。肉鰭魚類就完全有了這些特徵。牠們的顎骨結構及齒列與早期四足類是一致的。肉鰭魚類的一對鰭則較四足類的細小,但骨骼結構則很相似,有單一的近端骨頭、下一節有二塊骨頭,鰭細分成不規則的形狀,彷如手的腕骨或踝骨及指骨的結構。
早期四足類都是生存在石炭紀,牠們的祖先可能生存在更早的泥盆紀時期。早期的離片椎類都布著一些魚石螈的特徵,如相似的頭顱骨形態、迷齒列、頭顱骨上的關節、鰓結構及脊柱。但是牠們卻失去了幾種魚類的特徵,如尾巴上的輻鰭。
為了能在陸地上行走,四足類需要額外的支撐身體,因為在水中的浮力已經不存在。牠們亦須運用新的呼吸方法,直接從大氣中吸取氧氣,而非已溶於水中的氧。牠們亦須建立另一種運動模式。保存身體水份亦是一個重要的課題,因為離開水面後水份會大量及容易流失。新的感官系統亦需要來適應陸地的生活。
頭顱骨
四足類頭顱骨的特點在於前後部分的長度比例。魚類的後部較長,而眼窩在向前端位置。四足類頭顱骨的前部較長,眼窩在頭顱骨較後位置。淚骨並不再連接前部,且被前額骨分隔。頭顱骨亦可以自由的兩側移動。
離片椎類的後頂骨及顳骨之間有縫間關節,使扁骨與左右頂骨分隔。在頭顱骨後部,所有背部至匙骨的骨頭都消失了。
例如引螈的下顎,就像其肉鰭魚類的祖先般,外側有很長的齒骨。在齒骨與顎骨之間亦有骨頭,包括兩根夾骨、隅骨及上隅骨。內側一般都是三根鳥喙骨,有著牙齒及接近齒骨。在上顎中有一列迷齒,位於上頜骨及前上頜骨之上。引螈的牙齒會一浪一浪的更替,由最前排至最後排,每一顆成熟牙齒間必然是一顆新牙齒。
齒列
迷齒螈有獨特的牙齒結構,雖然並非只有牠們擁有如此的結構,但憑齒列就可以用來作出分類。牙齒上最特別的就是琺瑯質及象牙質形成縐狀,可以加強牙齒的結構及減低磨損。這些牙齒可以保存超過1億年,由肉鰭魚類帶到爬行動物。現今的兩棲類已沒有這種牙齒,取而代之的是連骨牙齒。
感官器官
四足類的感官器官若仍舊是與水中生活時一樣,在進入陸地後會對接收及反應化學訊號有一定難度。魚類有體側線來偵測水波動時的壓力。但是空氣的壓力卻不能偵測,但在迷齒螈的頭顱骨上有放置體側線感官器官的坑,可見牠們是半水中生活的。現今的兩棲類都是半水中生活的,其嗅覺上皮組織若須偵測空氣中的氣味,必須有一定程度的修改。
除了體側線外,眼睛亦需要改變。這是因空氣及水的折射率不同,故晶状体的焦距會有所改變。眼睛亦因暴露在較乾旱的環境,故需要發展眼皮及淚器來分泌液體滋潤眼睛。
聽覺
中耳保持平衡的功能是從魚類祖先承襲的,但空氣震盪卻不足以在頭顱骨造成脈衝,產生聽覺。大部分迷齒螈的噴水孔鰓腔都保留下來,成為耳裂,並由一塊薄及密的鼓膜所覆蓋。
魚類的舌骨下頜弓從顎骨的支撐位置往上移,縮小了形成鐙骨。鐙骨位於鼓膜及腦顱之間的空間,可以將頭顱外側的震盪帶到內部。故此鐙骨是阻抗匹配系統中的一個重要元素,將空氣傳播的聲波,帶到內耳的接收器中。這套系統是在不同兩棲類分支中獨立演化出來的。
為了令阻抗匹配的耳朵運作,鐙骨必須與鼓膜互相垂直,足夠細小及輕巧來減低慣性,並懸掛在充滿空氣的空間中。在現今對頻率超過1000赫茲的物種中,鐙骨底部是鼓膜面積的二十分之一。但是,早期兩棲類的鐙骨太大,阻礙了對高頻的聽覺。所以牠們可能只能聽到高強度及低頻的聲音,而鐙骨可能用來支撐腦顱的。[8]
骨骼
早期四足類的胸帶很大及有強壯的肌肉連接至肢體,可見是高度發展的。值得留意的是肩帶是與頭顱骨分離的,改進了陸地上的運動。肉鰭魚類的匙骨變成了鎖骨,間鎖骨位於胸部以下。在較為原始的四足類形態,兩個鎖骨及間鎖骨可能會向腹側生長,以形成較闊的胸板,但在引螈中卻不是這樣。胸帶的上部有肩胛骨,其下有可以作為肱骨連接的肩胛盂,向腹側有大而扁平,向著中線的鳥喙板。
現今的四足類的脊骨連接帶連接帶,而帶亦連接四肢,形成骨架。
骨盆亦較魚類為大,可以容納更多肌肉。骨盆向背部伸延,在脊骨位置以薦肋連合。後肢不只用來支撐體重,更可以行走。骨盆向背部伸延部分是髂骨,而闊的腹板則包含了前面的盆骨及後面的坐骨。這三根骨頭在骨盆的髖臼相連在一點,提供大腿骨的連接。
引螈的髂骨與薦骨是以韌帶連接,在結構上介乎最原始兩棲類及早期爬行動物。在較高等的脊椎動物都可見軟骨及薦肋都適合韌帶連接。
四肢
肱骨是手臂上最大的骨頭,扣在胸帶的髖臼內,遠端為橈骨及尺骨。橈骨是在前臂的內側,直接在肱骨之下,支撐著大部分的重量,而尺骨則在肱骨的外側。尺骨鷹嘴伸延到肱骨邊之上。
橈骨及尺骨與腕骨連接,腕骨共有三個部分,在橈骨底下的是橈側腕骨,尺骨底下的是尺側腕骨,並在兩者之間的中間物質。每根指頭相對都有一串五塊的遠端腕骨。每指的第一節稱為掌骨是在手掌位置。
骨盆肢骨與胸肢骨構造相同,但有不同名稱。與肱骨相對的是較長及幼的大腿骨。相對於橈骨及尺骨的是脛骨及腓骨,脛骨是在最內側,而腓骨則在最外側。踝骨則像腕骨。
攝食
早期四足類是有一個闊大的顎,以較弱的肌肉來開合。在顎內有像尖的上顎齒,配合張開的口,可見都是慣性的攝食習性。當兩棲類捕捉到獵物後,由於缺乏咀嚼的機能,會將擺動頭部,讓食物入到口的深處。這種攝食方法在今天的鱷魚及短吻鱷仍然可見。
現今兩棲類成體的舌頭是肉質的及與下顎前部連接,相信原始形態的亦是這樣連接,但卻不像蛙般專門。
早期四足類並不怎麼活躍,故此相信牠們不是獵食的。有可能是吃水中的魚類,或是在湖邊及沼澤的生物。加上當時非常豐富的無脊椎動物,估計有很足夠的食物供應。
呼吸
四足动物的肺是由肉鳍鱼的鱼鳔演化而来,通过横隔膜和肋间肌在胸腔内产生负压將空氣吸入肺泡,並通过毛细血管进行气体交换吸收氧气进行呼吸。兩棲類亦會經濕潤的皮膚及口腔黏膜來进行呼吸。引螈亦會呼吸,但由於其肋骨太近的關係,估計牠們會擴張胸籃。牠們可能張開嘴及鼻孔,壓迫舌骨來張開口腔,最終閉合嘴及鼻孔,並將空氣壓入肺部。從另一角度看,牠是先含著再吞下的。牠有可能透過肺壁上組織的收縮來呼氣。另外亦可能有其他特別的呼吸方法。
參考文獻
- Alexander, Pyron R. . Systematic Biology. 2011, 60 (4): 466–481 [2019-09-08]. PMID 21540408. doi:10.1093/sysbio/syr047. (原始内容存档于2020-01-31).
- Sues, Hans-Dieter. . Journal of Vertebrate Paleontology. 2019, 39: e1564758. doi:10.1080/02724634.2019.1564758.
- Di-Poï, Nicolas; Montoya-Burgos, Juan I.; Miller, Hilary; et al. . Nature. 2010-03-04, 464 (7285): 99–103 [2023-07-06]. PMID 20203609. S2CID 205219752. doi:10.1038/nature08789. (原始内容存档于2023-07-05).
- The World Conservation Union. 2014. IUCN Red List of Threatened Species, 2014.3. Summary Statistics for Globally Threatened Species. Table 1: Numbers of threatened species by major groups of organisms (1996–2014) (页面存档备份,存于).
- Vlad Tarko. . Sofipedia. [2008-02-18]. (原始内容存档于2008-02-22).
- . [2008-02-19]. (原始内容存档于2008-02-15).
- Swartz, B. . PLOS ONE. 2012, 7 (3): e33683. Bibcode:2012PLoSO...733683S. PMC 3308997
 . PMID 22448265. doi:10.1371/journal.pone.0033683.
. PMID 22448265. doi:10.1371/journal.pone.0033683. - . [2008-02-18]. (原始内容存档于2008-02-22).
延伸閱讀
| 维基共享资源上的相关多媒体资源:四足類 |
| 維基物種上的相關:四足類 |
- Benton, Michael. 3. John Wiley & Sons. 5 February 2009: 1 [10 June 2015]. ISBN 978-1-4051-4449-0. (原始内容存档于2020-08-20).
- Clack, J.A. 2nd. Bloomington, Indiana, USA.: Indiana University Press. 2012. ISBN 9780253356758.
- Laurin, Michel. . University of California Press. 2010 [26 May 2015]. ISBN 978-0-520-26647-6. (原始内容存档于2020-08-19).
- McGhee, George R. Jr. . Columbia University Press. 2013 [2 May 2015]. ISBN 978-0-231-16057-5. (原始内容存档于2020-08-20).
- Steyer, Sebastien. . Indiana University Press. 2012: 59 [1 June 2015]. ISBN 978-0-253-22380-7. (原始内容存档于2020-09-17).
- Clack, Jennifer A. . Annual Review of Earth and Planetary Sciences. 2009, 37 (1): 163–179. Bibcode:2009AREPS..37..163C. doi:10.1146/annurev.earth.36.031207.124146.
- Hall, Brian K. (编). . Chicago: University of Chicago Press. 2007 [2020-09-06]. ISBN 978-0-226-31340-5. (原始内容存档于2020-08-01).
- Long JA, Young GC, Holland T, Senden TJ, Fitzgerald EM. . Nature. November 2006, 444 (7116): 199–202. Bibcode:2006Natur.444..199L. PMID 17051154. doi:10.1038/nature05243.
- Benton, Michael. 3rd. Blackwell Publishing. 2005.