基底核
基底核(英語:)又称基底节、基底神经节,是大脑深部一系列神经核团组成的功能整体。它是位於大腦皮質底下一群運動神經核的統稱,[1]与大脑皮层,丘脑和脑干相连。目前所知其主要功能为自主运动的控制、整合調節細緻的意識活動和運動反應。它同时还参与记忆,情感和奖励学习等高级认知功能。基底核的病变可导致多种运动和认知障碍,包括帕金森氏症和亨廷顿氏症等。
| 基底核 | |
|---|---|
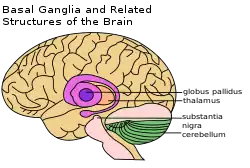 右上方有標示出基底核 | |
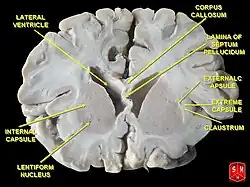 從腦的冠狀切面觀察基底核 | |
| 基本 | |
| 屬於 | 大腦 |
| 标识字符 | |
| 拉丁文 | nuclei basales |
| MeSH | D001479 |
| NeuroNames | 224, 2677 |
| NeuroLex ID | birnlex_826 |
| TA98 | A14.1.09.501 |
| TA2 | 5559 |
| FMA | FMA:84013 |
| 《神经解剖学术语》 | |
命名
基底核的英文学名为“basal ganglia”,basal(基底)意表起解剖位置位于大脑的深部和底部,而 ganglia(神经节)是一命名错误。Ganglia 一词本义指周邊神經系統的神经元胞体的聚集部,但实际上基底核位于中枢神经系统。中枢神经系统的神经元胞体聚集应称为“核”(nucleus),例如红核(red nucleus)。这个英文的错误命名一直沿用至今,成为习惯[2]。但是中文术语“基底核”却纠正了这个错误。
研究历史
英国解剖学家托马斯·威利斯(Thomas Willis)于1664年最早记载了基底核的结构。 [3] 之后许多年,“纹状体”(Corpus striatum)一词被用来泛指皮层下的一系列核团。 但是后来发现,其中许多核团之间并没有直接的联系。比如壳核(Putamen)和尾状核(Caudate nucleus)之间就没有直接的连接。 今日的“纹状体”(Striatum)一词指尾状核,壳核和伏隔核三者构成的整体,该整体是基底核系统的主要输入通道。该命名法则是Cécile and Oskar Vogt (1941) 提出的。 纹状体得名于其多纹的外形。该外形来自于大量致密的纹状体-苍白球-黑质(纹苍黑)纤维。这些纤维连接构成了基底核神经回路的核心。纹苍黑纤维在跨过内囊的部分也叫做“Edinger梳状束”。 苍白球(Globus pallidus)在早些时候被称为豆状核(Lentiform nucleus)。
Luys于1865年发现了丘脑下核(Subthalamic nucleus)。晚些时候,人们发现大脑脚-脑桥复合体(Pedunculopontine complex)和中央复合体(Central complex)可能调控基底核的功能。20世纪的医学和神经生物学研究并且从初步从机理上认识了基底核的损伤和一些列运动疾病(例如帕金森氏症和亨廷顿氏症)的关联,并且澄清了基底核参与运动和认知功能。
解剖构造
如图所示,灵长目的基底核的主要组成部分如下:
前侧
- 纹状体(Striatum)包括
后侧,以下这些结构在大脑中更靠下,靠后。
- 丘脑下核(Subthalamic nucleus, STN)
- 黑质(Substantia nigra, SN),根据内部结构可分为
- 黑质致密部(Substantia nigra pars compacta,SNc)
- 黑质网状部(Substantia nigra pars reticulata,SNr)
- 黑质侧部(Substantia nigra pars lateralis,SNl)
应注意不同学者经常赋予基底核不同的范围。例如,并非所有学者主张将黑质算作基底核的一部分。也有一些学者主张将杏仁核划入基底核。 哺乳类的大脑包含左右两侧的一对基底核,关于大脑中线对称。 基底核可以从功能上划分为一系列并行回路,包括骨骼肌肉运动回路,边缘回路和动眼回路等。不同的回路投射到不同的目标区。例如骨骼肌肉运动回路主要投射到丘脑的腹外侧核(Ventrolateral nucleus, VL)。
回路和神经递质
左图所示为基底核的基本内部连接。纹状体(包括尾状核和壳)是基底核系统的主要输入通道。大脑皮层的诸多运动,前额叶和感觉分区投射到纹状体。如表2所示,基底核中不同核团的神经元合成不同的神经递质。谷氨酸是一种兴奋性神经递质,GABA是一种抑制性神经递质。而如下表所示,多巴胺的兴奋或抑制取决于受体的种类。来自黑质致密部的多巴胺能输入刺激纹状体内的所有多巴胺受体,但是由于间接和直接通路内的受体种类不同,多巴胺输入的效应也不同。从下表中可以看出,由于多巴胺输入对直接通路的效应是兴奋,而对间接通路的效应是抑制,该神经递质的总体功能是激活丘脑。 [4]
基底核包含两大回路,分别成为直接通路(Direct pathway)和间接通路(Indirect pathway)。它们是皮质-基底核-丘脑回路(Cortico-basal-thalamic loop)的两大部分。
表1,基底核的直接与间接通路
| 通路名称 | 通路的组成 (+表兴奋性输出, -表抑制性输出 | 抑制性连接的个数 | 机能描述 | 多巴胺受体类型 |
| 直接 | 大脑皮质+ →纹状体- →内苍白球/黑质(GPi/SNr)- →丘脑+ →大脑皮质 | 2 (偶) | 大脑皮质的活动激活纹状体神经元,进而抑制内苍白球的自发发放。由于苍白球的神经元对丘脑的作用是抑制性的,这一效应实际上削弱了丘脑所受到的抑制性输入。这一过程成为“去抑制”(Disinhibition) | D1 (兴奋性) |
| 间接 | 大脑皮质+ →纹状体- →外苍白球(GPe) - →丘脑下核(STN) + →内苍白球/黑质(GPi/SNr)- →丘脑+ →大脑皮质 | 3 (奇) | 与直接通路不同,由于间接通路中抑制性突触的个数为奇数,纹状体神经元细胞的激活对丘脑产生抑制效应。 | D2 (抑制性) |
表2,基底核的神经递质
| 核团 | 神经递质 | 描述 | 相关疾病 |
| 纹状体 | GABA | 纹状体的输出神经元是GABA能,抑制性的。 | 亨廷顿氏症。 |
| 黑质 | 多巴胺 | 黑质致密部的神经元向纹状体发出多巴胺能投射。其效应取决于受体种类。 | 若黑质致密部细胞的多巴胺合成受阻,纹状体受到的多巴胺能输入的降低会导致运动和认知障碍。最典型的例子是帕金森氏症。 |
| 苍白球 | GABA | 苍白球包括内外两大部分。内苍白球直接投射到丘脑,而外苍白球直接投射到丘脑下核。两者的作用都是抑制性的。 | 妥瑞氏综合征(Tourette's syndrome),亦称抽动秽语综合征。 |
| 丘脑下核 | 谷氨酸 | 丘脑下核的神经元通过谷氨酸能投射来兴奋内苍白球细胞。这是间接通路的一部分 |
丘脑下核的损伤造成半侧投掷症(hemiballismus) [5] |
细胞类型和突触连接
人类的纹状体中大约有一亿神经元。70%以上为输出神经元。这些输出神经元在其树突上分布有非常致密的棘头(Spine),故称为棘状神经元。 这些棘头是棘状神经元与传入纹状体的轴突形成突触的部位。输入纹状体的轴突主要来自大脑皮层,丘脑和杏仁体。 这些轴突进入纹状体后,展出覆盖范围甚大的侧枝,与大量纹状体神经元形成突触。反过来,每个纹状体神经元也和大量的输入轴突形成突触。据估计,每个棘状神经元接受来自7500-15000个不同轴突的输入。 这种连接模式类似于小脑的平行纤维和普肯野细胞(Purkinje cells)的连接模式。它可能类似于一种电子电路中的组合逻辑。 输入到纹状体棘状神经元的纤维(即轴突)大多为兴奋性的。但是这些神经元大多数时候却处在静息状态(即不发放动作电位)的状态。 棘状神经元的这种活动模式,如果考虑到其收到的兴奋性输入的个数,则相当难以理解。其机制目前尚在研究中。 [6]
纹状体中其余的神经元为中间神经元(Interneurons),这些神经元都为抑制性,包括胆碱能神经元,GABA/Parvalbumin神经元和Somatostasin/NOS神经元等。这些抑制性中线神经元也接受来自大脑皮质,丘脑和杏仁体的输入,并且和纹状体的棘状神经元相互形成突触连接。
在人类及其他灵长目动物中,纹状体中有明显的“镶嵌”(Mosaic)结构。在该镶嵌结构中,纹状体内有神经元密度较高的区域,称为“纹状小体”(Striasome),以及将这些纹状小体隔开的“基质”(Matrix)。纹状体的传入纤维在很大程度上遵守基质所划定的界限。而且纹状小体和基质中的神经元似乎也投射到黑质的不同部位。所以纹状小体和基质似乎在纹状体中形成了两个并行的通路,其功能重要性尚在研究中。该并行通路不应与前面所讲的直接和间接通路混淆。[6]
纹状体棘状神经元向苍白球和黑质发出GABA能抑制性输出。据估计,苍白球大约只有70万神经元,比纹状体神经元的数量少两个数量级。这就意味着平均来说,每个苍白球神经元接受来自100个左右纹状体神经元的抑制性输入。
苍白球和黑质的输出神经元在大多数时候处在自发发放状态,这可能与这些细胞的膜特性有关。它们投射到丘脑腹外侧核以及上丘,其自发持续发放抑制其中的细胞。但是一旦纹状体内的特定棘状神经元被激活,这些苍白球/黑质神经元将受到强有力的抑制,从而暂停发放。这一过程释放了苍白球对丘脑腹外核的抑制,使一个动作得以启动。
與基底核功能相關的疾病 : 基底核的病變可導致多種運動和認知障礙,如 帕金森氏症、亨丁頓舞蹈症、半側投擲症、妥瑞氏症候群、肌肉張力失常、 痙攣性失聲症、口吃。
学者已经在猴子等动物进行了一系列在体(In vivo)电生理研究。在动物体内,棘状神经元多数时候静息,只偶尔发放若干个脉冲,且持续时间不超过3秒,发放率不超过40脉冲/秒。这些脉冲一般与动物的特定动作的起始同步。另一些实验观察到了动物在启动响应性动作(例如动眼)时黑质细胞的暂停发放 [7] [8]。 这种对应于特定感觉指导下的动作的基底核神经元发放模式是随着动作的学习逐渐产生的 [9]。 另外,有趣的是,一项研究表明:许多黑质细胞自发发放的暂停,只发生在依赖与外界感觉信号的,学习到的动作,而不发生在自发的动作,即使这两个是同一个动作 [10]。
功能
眼球运动
基底神经节的一项受到深入研究的功能是它在控制眼球运动中的作用。[11]眼球运动受到广泛的大脑区域网络的影响,该网络集中在称为上丘的中脑区域。上丘是一种分层结构,其层形成视觉空间的二维视网膜图。上丘深层神经活动的“碰撞”驱动眼球运动指向空间中的相应点。
上丘接收来自基底神经节的强烈抑制性投射,起源于黑质网状部。黑质网状部中的神经元通常以高速率连续发射,但在眼球运动开始时它们“暂停”,从而使上丘摆脱抑制。所有类型的眼球运动都与信噪比中的“暂停”有关;然而,单个黑质网状部神经元可能与某些类型的运动比其他类型的运动更密切相关。尾状核某些部分的神经元也表现出与眼球运动相关的活动。由于绝大多数尾状细胞以非常低的速率发射,因此这种活动几乎总是表现为发射速率的增加。因此,眼球运动从尾状核的激活开始,尾状核通过直接的 GABA 能投射抑制信噪比,进而抑制上丘。
在积极性中的作用
基底神经节中的细胞外多巴胺与啮齿动物的积极性有关,高水平与饱足的“欣快感”有关,中等水平与寻求有关,低水平与厌恶有关。边缘基底神经节回路受到细胞外多巴胺的严重影响。增加的多巴胺导致腹侧苍白球、内足核和黑质网状部的抑制,导致丘脑去抑制。这种直接 D1 和间接 D2 通路的模型解释了为什么每个受体的选择性激动剂没有回报,因为去抑制需要两种通路的活性。丘脑的去抑制导致前额叶皮层和腹侧纹状体的激活,选择性地增加 D1 活动导致奖励。[12]还有来自非人类灵长类动物和人类电生理学研究的证据表明,包括苍白球内部和丘脑底核在内的其他基底神经节结构也参与了奖赏处理。[13][14]
做决定
研究人员已经为基底神经节提出了两种模型,一种是由腹侧纹状体中的“批评者”产生并估计值的动作,以及由背侧纹状体中的“执行者”执行的动作。另一种模型提出基底神经节作为一种选择机制,其中动作在皮层中生成,并由基底神经节根据上下文进行选择。[15]CBGTC 循环也涉及奖励折扣,随着意外或高于预期的奖励而增加。[16]一篇综述支持这样一种观点,即无论结果如何,皮层都参与学习行动,而基底神经节则参与基于基于联想奖励的试错学习来选择适当的行动。[17]
工作记忆
有人提出,基底神经节可以控制进入和不进入工作记忆的内容。一个假说提出直接通路(Go 或兴奋性)允许信息进入前额皮质,在那里它保持独立于通路;另一种理论认为,为了使信息留在前额皮质中,直接通路需要继续反射。有人提出,短的间接通路是与直接通路相拮抗的,可以关闭通向前额皮质的门。这些机制共同调节工作记忆焦点。[18]
与基底核功能相关的疾病
如上文所述,帕金森氏症,表现为运动迟缓,运动丧失和静止性震颤等症状,主要是由于黑质致密部的退化和随后的多巴胺能输出降低有关。多巴胺的减少造成了纹状体直接通路的抑制和间接通路的去抑制,进而导致基底核对丘脑的兴奋的丧失。与帕金森氏症几乎相对的一种疾病成为亨廷顿氏症(Huntington's disease),表现为异常的多餘动作(舞蹈性震颤)以及协调缺乏等症状,是由于纹状体和苍白球的退化造成的。半侧投掷症(Hemiballism),表现为反覆的上肢运动(如投掷棒球),是由于丘脑下核(subthalamus)的退化或损伤。丘脑下核作为间接通路的最後一级,其到兴奋内苍白球的作用。这种兴奋的确实导致内苍白球对丘脑的抑制作用减弱,进而导致了半侧投掷症中所见的冗餘动作(多动症)[5]。妥瑞氏综合征,表现为运动痉挛,注意力问题和口出秽语等症状,也被认为与基底核的兴奋和抑制作用的平衡丧失有关 [19]。 目前被认为可能与基底核功能相关的疾病还包括张力失常(dystonia) [20] [21], 痉挛性失声症和口吃 [22] 等。
参考文献
- 成人內外科護理學
- Soltanzadeh, Akbar. . Tehran: Jafari. 2004. ISBN 964-6088-03-1.
- Andrew Gilies, A brief history of the basal ganglia (页面存档备份,存于), retrieved on 27 June 2005
- Delong, M. . E. R. Kandel, J. H. Schwartz, & T. M. Jessell (编). 4th edition. New York: McGraw-Hill. 2000.
- Bear, Mark F.; Barry W. Connors and Michael A. Paradiso. 2nd edition. Lippincott Williams & Wilkins Inc. 2001: 480–481.
- Wilson, C. J. . Sheperd, G. M. (编). 4th edition. New York: Oxford Univ. Press. 1998.
- DeLong, M. R. . Science. 1973, 179: 1240–1242.
- Kimura, M., Raijkowski, J., & Evarts, E. . Proc. Natl. Acad. Sci. USA. 1984, 81: 4998–5001.
- Graybiel, A. M., Aosaki, T., Flaherty, A. W., & Kimura, M. . Science. 1994, 265: 1826–1831.
- Hikosaka, O, & Wurtz, R. H. . Brain Res. 1983, 272: 368–372.
- Hikosaka, O; Takikawa, Y; Kawagoe, R. . Physiological Reviews. 2000, 80 (3): 953–78. PMID 10893428. S2CID 7502211. doi:10.1152/physrev.2000.80.3.953.
- Eisinger, Robert S.; Urdaneta, Morgan E.; Foote, Kelly D.; Okun, Michael S.; Gunduz, Aysegul. . Frontiers in Neuroscience. 2018, 12: 385. ISSN 1662-453X. PMC 6041403
 . PMID 30026679. doi:10.3389/fnins.2018.00385 (英语).
. PMID 30026679. doi:10.3389/fnins.2018.00385 (英语). - Eisinger, RS; Urdaneta, ME; Foote, KD; Okun, MS; Gunduz, A. . Frontiers in Neuroscience. 2018, 12: 385. PMC 6041403 . PMID 30026679. doi:10.3389/fnins.2018.00385 .
- Redgrave, P.; Prescott, T.J.; Gurney, K. (PDF). Neuroscience. April 1999, 89 (4): 1009–1023 [23 September 2019]. PMID 10362291. S2CID 3187928. doi:10.1016/S0306-4522(98)00319-4. (原始内容存档 (PDF)于25 July 2018).
- Maia, Tiago V.; Frank, Michael J. . Nature Neuroscience. 15 January 2017, 14 (2): 154–162. ISSN 1097-6256. PMC 4408000 . PMID 21270784. doi:10.1038/nn.2723.
- Hélie, Sébastien; Ell, Shawn W.; Ashby, F. Gregory. . Cortex. 1 March 2015, 64: 123–135. ISSN 1973-8102. PMID 25461713. S2CID 17994331. doi:10.1016/j.cortex.2014.10.011.
- Scahill, L., Erenberg, G., Berlin, C. M. Jr., Budman, C., Coffey, B. J., Jankovic, J., Kiessling, L., King, R. A., Kurlan, R., Lang, A., Mink, J., Murphy, T., Zinner, S., & Walkup, J. . NeuroRx. 2006, 3(2): 192–206. PMID 16554257
- Dystonia fact sheet: National Institute of Neurological Disorders and Stroke . [2007-01-03]. (原始内容存档于2006-12-05).
- Blood, A. J., Tuch, D. S., Markis, N., Makhlouf, M., Sudarsky, L. R., Sharma, N. . Neuroreport. 2006, 17: 1251–1255.
- Alm PA. (PDF). Journal of communication disorders. 2004, 37 (4): 325–69. PMID 15159193.