恐爪龍屬
恐爪龍屬(屬名:)是馳龍科恐龍的一屬,身長約2.5-4公尺,生活於下白堊紀的阿普第階中期至阿爾布階早期,距今約1億1500萬至1億800萬年前。牠的學名是來自古希臘文的「δεινος」(意為「恐怖」)及「ονυξ/ονυχος」(意為「爪」),因為牠的後肢第二趾上有非常大,呈鐮刀狀的趾爪,在行走時第二趾可能會縮起,僅使用第三、第四趾行走。一般認為恐爪龍會用其鐮刀爪來割傷獵物,但近年就伶盜龍重建模型的測試顯示這爪作刺戳之用,而非割劃。
| 恐爪龙属 | |
|---|---|
 | |
| 恐爪龍的骨架模型,位於菲爾德自然史博物館 | |
 | |
| 平衡恐爪龍的重建圖 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 亚目: | 獸腳亞目 Theropoda |
| 科: | †驰龙科 Dromaeosauridae |
| 演化支: | †真驰龙类 Eudromaeosauria |
| 亚科: | †驰龙亚科 Dromaeosaurinae |
| 属: | †恐爪龙属 Deinonychus Ostrom, 1969 |
| 模式種 | |
| †平衡恐爪龍 Deinonychus antirrhopus Ostrom, 1969 | |
| 異名 | |
| |
恐爪龍的正模標本(編號YPM 5205)有個大幅彎曲的第二趾爪,奧斯特倫姆研究鱷魚、鳥類的趾爪,發現主龍類的趾爪具有角質鞘,所以趾爪的實際體積較大。奧斯特倫姆估計這個第二趾爪有13公分長[1]。
模式種為平衡恐爪龍(D. antirrhopus),種名antirrhopus意為「平衡」,取自於奧斯特倫姆對於尾巴功能的假設。就像其他的馳龍科,恐爪龍的尾巴有者一連串的長骨突與骨化肌腱。這種結構使恐爪龍的尾巴硬挺,可提供更好的平衡及轉彎能力。但是一個蒙古伶盜龍的標本(編號IGM 100/986),尾巴是往後彎曲、呈S狀,而且關節未脫落。這個發現顯示尾巴可以側彎,並有一定的靈活性[2]。
古生物學家約翰·奧斯特倫姆(John Ostrom)在1960年代末期的恐爪龍研究有突破性的影響,引發恐龍究竟是否為恆溫動物的爭論,這個改變稱為恐龍文藝復興。在這之前,一般大眾認為恐龍是巨大、行動緩慢的爬行動物。奧斯特倫姆根據恐爪龍的小型體型、類似鴕鳥的二足步態、與地面平行的尾巴,以及大型、鐮刀狀的第二趾爪,提出恐爪龍是靈活、行動敏捷的獵食動物[3]。
恐爪龍的化石大多數發現於美國蒙大拿州、懷俄明州的Cloverly組,以及奧克拉荷馬州的鹿角組。而在馬利蘭州大西洋沿岸平原波多馬克組發現的牙齒,也可能屬於恐爪龍。在Cloverly組及鹿角組發現的恐爪龍化石與鳥腳下目腱龍的化石位置接近。根據與腱龍化石一同被發現的牙齒標本,顯示這些恐爪龍正在獵食這隻腱龍,或是恐爪龍正在食用腱龍的屍體。另外恐爪龍因侏羅紀公園系列電影的緣故,其體型被誤會是迅猛龍的體型。
描述
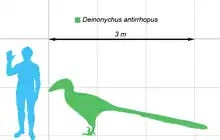
根據恐爪龍的最大型標本,恐爪龍的體型可達3.5公尺,頭顱骨最大可達41公分長,臀部高度為0.87公尺,而體重可达90公斤[4]。而恐爪龙属中最大的个体(不一定属于平衡恐爪龙),YPM 5210 更是可以达到长4公尺,重120公斤。牠的頭顱骨有強壯的頜部,有約60根彎曲、刀刃形的牙齒。奧斯特倫姆最初將恐爪龍的不完全顱骨,重建成三角形、寬廣的頭部,類似異特龍。在發現更多、更完整的恐爪龍與其近親的化石後,顯示恐爪龍的上齶部較呈拱形,口鼻部較狹窄,顴骨寬廣,使頭部看起來較為立體,與奧斯特倫姆的重建不同[5]。恐爪龍顱骨與伶盜龍不同,頭頂較堅固,類似馳龍;伶盜龍的鼻部上側往下凹,這點與恐爪龍不同[6]。頭顱骨及下頜都有洞孔,可減輕頭部重量,而恐爪龍的眶前孔是特別的大[5]。按頭顱骨來推算,眼睛主要是向兩側的。
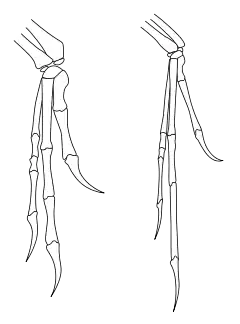
就像其他的馳龍科,恐爪龍有大型手掌與三根手指。第一指最短,而第二指最長。每隻後肢的第二趾都有鐮刀狀的趾爪,長度約12公分,有可能是作為捕獵動物。牠可以先向前戳刺,並向下割來撕破獵物。相對於恐爪龍的體型,這些趾爪來的相當地大。恐爪龍的身體是靠尾椎及人字骨,在高速轉向時來維持穩定及平衡。
分類
恐爪龍是最著名的馳龍科恐龍之一[7],且是伶盜龍的近親,伶盜龍的體型較小,生存於上白堊紀的蒙古[8][9]。不过牠們的不属于同一亚科。恐爪龙属于驰龙亚科。而伶盗龙属于伶盜龍亞科,這個亞科最初是由瑞欽·巴思缽(Rinchen Barsbold)於1983年所提出[10],而當時只有單一的伶盜龍屬。与伶盜龍亞科纤细的头骨不同,驰龙亚科的头骨更为宽厚,坚硬,可以发挥出巨大的咬合力。馳龍科與傷齒龍科共同組成恐爪龍下目,是鳥類的姊妹分類單元。在種系發生學上,恐爪龍下目代表了一群不屬於鳥類,卻與鳥類是近親的恐龍[11]。
發現與命名


恐爪龍的化石發現於美國蒙大拿州與懷俄明州的Cloverly組[12],以及奧克拉荷馬州的鹿角組[13]。Cloverly組的地質年代為白堊紀早期的阿普第階晚期至阿爾布階早期,距今約1億1500萬到1億800萬年前[14][15]。此外,馬里蘭州大西洋沿岸平原地帶的波多馬克組,發現了一些可能屬於恐爪龍的牙齒,年代為阿普第階[16]。
恐爪龍的第一副化石,是在1931年由巴納姆·布郎(Barnum Brown)所帶領的隊伍在蒙大拿州南部的Cloverly組發現。布郎當時主要是想發掘並處理腱龍的遺骸,但在他交予美國自然史博物館的1931年報告中,指出發現了一小型的肉食性恐龍,發現位置接近腱龍化石,但因陷在石灰岩而難以作清潔處理[17]。他非正式地將之命名為"Daptosaurus",並準備描述及展示其骨骼,但他卻沒有完成[18]。另外,布朗將一個恐爪龍的牙齒,與另一種較小型獸腳類恐龍的骨骸混淆在一起,並非正式命名為"Megadontosaurus"。
約30年後,從1964年8月開始,古生物學家約翰·奧斯特倫姆(John Ostrom)率領一個耶魯大學皮博迪自然史博物館的挖掘團隊,在其後的兩年內發現了超過1000個骨頭,來自於至少三個個體。因為很難判斷這些化石的正確位置,所以恐爪龍的正模標本(編號YPM 5205)只限於完整的左足部,與部分的右足部,但都確定屬於同一個體[19]。其他標本則在耶魯大學的皮博迪自然史博物館區分、標示成50個不同的項目。
在接下來幾年,奧斯特倫姆及格蘭特·邁耶(Grant E. Meyer)研究這些新發現化石,以及布朗所命名的"Daptosaurus"化石,發現牠們是同種生物,並在1969年將這些化石命名為平衡恐爪龍(Deinonychus antirrhopus)。種名antirrhopus意為「平衡」,指的是牠們堅挺尾巴的可能用途[20]。同年,奧斯特倫姆公布更廣泛的恐爪龍專題論文[19]。
奧斯特倫姆同時重新檢驗布朗所命名的"Megadontosauru",他發現那隻牙齒是屬於恐爪龍,而骨骼則是來自另一類動物。他在1970年,將骨骼部分命名為小獵龍[18]。
雖然在1969年發現大量的恐爪龍化石,但許多重要部位的骨頭並沒有發現,剩下的難以確認。這些化石包含少數眼眶後顱骨,沒有股骨、沒有薦骨、沒有叉骨或胸骨、也沒有脊椎,以及一個被奧斯特倫姆認為是鳥喙骨的小型碎片。奧斯特倫姆將恐爪龍重建成具有獨特的骨盆,恥骨呈梯形且平坦,與坐骨長度一樣。
更多發現

在1974年,奧斯特倫姆公佈了另一份恐爪龍專題論文,並指出他原本鑑定的恥骨,其實是個肩膀上的喙骨[21]。同一年,法里什·詹金斯(Farish Jenkins)所率領的哈佛大學挖掘團隊,在蒙大拿州發現了另一個恐爪龍標本。這個新標本增添了七個沒發現過的部位,包含:股骨、恥骨、薦骨、腸骨、以及腳掌與蹠骨。奧斯特倫姆研究了這個新標本,並推翻他之前的恐爪龍重建。這次恐爪龍被重建成具有長的恥骨,而奧斯特倫姆並推論恥骨稍微後傾,類似鳥類[22]。
在紐約的美國自然史博物館,有一個展覽中的恐爪龍骨架,上有最初的標本組成[23]。哈佛大學的比較動物博物館則有其他的標本展覽中。它們的恐爪龍骨架都缺乏胸骨、胸肋、叉骨、腹肋。耶魯大學也有恐爪龍骨架,但與前兩者的發現處不同。
在奧斯特倫姆研究了的正模標本後,仍有幾個陷在石灰岩的小型標本未被處理,而擺放在美國自然史博物館。這些標本大部份是獨立的骨頭及骨頭碎片,包括了標本埋葬時的周圍母岩。在2000年時,Gerald Grellet-Tinner與彼得·馬克維奇(Peter Makovicky)重新研究這些標本,發現一些有趣及被忽略的特徵。當中的幾條修長骨頭,原先被認為是骨化肌腱,可使恐爪龍尾巴堅硬,其實是腹肋。而且在原先恐爪龍標本的周圍,也發現了一些被忽略的蛋殼化石[24]
在後來的詳細報告中,Tinner與馬克維奇提出這些蛋幾乎肯定是屬於恐爪龍的,是首批發現的馳龍科蛋[17]。再者,其中一個蛋殼的外表面緊接恐爪龍的腹肋,可見恐爪龍可能正在孵蛋。由此推斷恐爪龍是用身體的熱量來孵蛋,類似現今鳥類,這也顯示牠們是溫血動物[25]。在2007年,格里高利·艾利克森(Gregory Erickson)與其同事的進一步研究,發現正模標本的年齡約13或14歲,而且是穩定的成長。同一份研究中也指出,恐爪龍在死亡前便已停止成長,與其他已知的獸腳類恐龍不同[26]。
影響
奧斯特倫姆在1969年的恐爪龍研究,被認為是20世紀中期最重要的恐龍發現[27]。因為這個新發現的活躍、敏捷恐龍,改變了很多關於恐龍的科學研究,以及一般大眾對恐龍的認知,且開啟了對恐龍可能是溫血動物的辯論。這個認知的變化稱為恐龍文藝復興。幾年後,奧斯特倫姆發現了恐爪龍與鳥類的手掌的相似性,使他認為鳥類是從恐龍所演化而來[28]。三十年後,這個觀念幾乎是被普遍地接受。
目前還沒有發現恐爪龍具有羽毛的證據。但某些馳龍科化石已發現有羽毛的直接與間接證據,顯示這個演化支普遍具有羽毛[29]。生存年代較早、較為原始的小盜龍,也屬於馳龍科[30],已有多個標本被發現具有羽毛痕跡,分布於前肢、腿部、尾巴,羽毛形態已經相當類似現代鳥類[29]。另一種在中國發現的中國鳥龍,化石也被發現有类似羽毛的結構,其特徵顯示牠們較恐爪龍更為原始,其生存年份也早於恐爪龍[31]。在2007年,恐爪龍的近親伶盜龍被發現前肢尺骨具有羽莖瘤。現代的鳥類也具有羽莖瘤,是羽毛的附著處。這個發現顯示伶盜龍也具有羽毛[32]。在其他獸腳亞目中,如尾羽龍、鳥面龍、北票龍及帝龍,也有發現羽毛的輪廓,根據親緣分支分類法的位置,更令人相信所有馳龍科都是有羽毛的[33]。菲力·森特(Phil Senter)在進行恐爪龍、斑比盜龍的前肢活動範圍研究時,提出馳龍類可能都具有飛羽[34]。
古生物學
棲息地
地質學證據顯示恐爪龍棲息於氾濫平原或沼澤[7]。Cloverly組及鹿角組的古生態環境包含有熱帶與副熱帶森林、三角洲及潟湖,並類似今日的路易斯安那州[35][36]。
其他同時期的恐龍包括有草食性的楯甲龍、鳥腳下目的西風龍及腱龍。在奧克拉荷馬州,恐爪龍的生態系統也包括了巨大獸腳亞目的高棘龙、蜥腳下目的波塞東龍、稜角鱗鱷、以及雀鱔目的Lepisosteus[36]。如果馬里蘭州所發現的牙齒是屬於恐爪龍的,那恐爪龍也在當地區與星牙龍、傷龍、孔牙龍(目前只發現牙齒)等恐龍共同生存。Cloverly組中段的地質年代約為1億1500萬[14]到1億800萬年前[15]。
獵食行為

在Cloverly組,常在腱龍化石的附近發現恐爪龍的牙齒。而在其中兩個採石場,已發現相當完整的恐爪龍化石,就位在腱龍化石的附近。第一個是蒙大拿州Cloverly組的耶魯採石場,發現過四個恐爪龍的成年個體與一個幼年個體,以及眾多的牙齒。基於在同一位點發現大量恐爪龍的骨骼,且在腱龍的附近發現恐爪龍的牙齒,估計恐爪龍是獵食腱龍的。奧斯特倫姆與德斯蒙德·麥斯威爾(W. Desmond Maxwell)更以此推斷恐爪龍是成群生活及獵食的[37]。第二個採石場則位在奧克拉荷馬州的鹿角組。該地發現六個腱龍的部份骨骸,體型不一,附近另有一個恐龍的部份骨骸與眾多牙齒。一個腱龍的肱骨上有齒痕,可能是恐爪龍所留下。在1998年,丹尼爾·布林曼(Daniel L. Brinkman)等人提出成年的恐爪龍為70到100公斤,而成年腱龍為1到4公噸。單一的恐爪龍無法獵殺成年的腱龍,顯示牠們應是採群體方式獵食腱龍[38]。
但是在2007年,布林曼與B. T. Roach基於現今肉食性動物的獵食方式及腱龍位點的埋葬學,質疑了恐爪龍在群獵時的合作性行為。現今的主龍類(鳥類及鱷魚)及科莫多龍有少數的合作性捕獵,不過牠們主要是單獨獵食,或是被吸引至已死的屍骸,而會為了搶食獵物屍體而發生打鬥。例如,當一群科莫多龍一同攝食時,最大的會先吃,且會攻擊其他嘗試吃食的較小型科莫多龍;如果小型的科莫多龍被殺死,牠們的屍體會被同類搶食。當這些資料套用在腱龍與恐爪龍的位點時,這似乎與科莫多龍與鱷魚的攝食模式很吻合。在這些位點發現的恐爪龍骨骼都是接近成年的,而失去了的部位可能是被其他恐爪龍吃了[39]。在2007年,科學家們在中國山東省發現了第一個大量的馳龍科足跡化石。這個足跡化石可能是由大型馳龍科恐龍所留下,而且是由六個相近大小的個體,沿者海岸共同前進。這些個體相距約一公尺,朝同一方向緩慢前進。研究人員提出,雖然該足跡化石並不能明確指出馳龍科的獵食行為,但無法排除牠們以群體獵食的可能性[40]。
.jpg.webp)

在2011年的一項恐爪龍下目趾爪研究,比對不同恐爪龍類、現代鳥類的腳掌結構、趾爪形狀,根據這些現代鳥類的獵食行為,分析這群恐龍的壓制獵物能力。這項研究認為伶盜龍等馳龍類可能以小型獵物為食。研究發現,許多掠食性現代鳥類也有大型第二趾爪,當牠們獵食小型獵物時,會使用腳掌、趾爪將獵物固定在地面上,用體重施壓於獵物,再用鳥喙吞食這些獵物,尤其是鷹科。這項研究推論馳龍類會以類似的方式捕抓、固定獵物,並開始吞食獵物的身體,直到獵物失血過多、失去生理機能。研究發現馳龍類的後肢、腳掌結構最類似鷹,也有大型第二趾爪、類似的腳掌抓取範圍;馳龍類的跗跖骨短、腳掌力量,則比較類似貓頭鷹。伶盜龍的頜部、前肢結構,也符合這項研究的結論。馳龍科的前肢強壯,但極可能覆蓋者羽毛,牠們吞食獵物時可能會將前肢張開,以伺機保持身體的平衡;尾巴也具有平衡身體的功能。馳龍科的頜部結構虛弱,可能主要用來吞食、撕咬獵物的肉體,而非以快速、強壯力量攻擊獵物。研究人員推測,近鳥類可能共同擁有這些演化特徵,而現代鳥類則繼承這些特徵[41]。
咬合力
在2005年,一群科學家重建恐爪龍的頜部肌肉群,計算恐爪龍的咬合力,發現牠們的咬合力相當於現代美洲鱷的約15%[42]。在2010年,另一群科學家研究一個腱龍化石上的恐爪龍齒痕,發現這個恐爪龍是個大型個體,而且能直接咬穿大型動物的骨頭。他們計算出的恐爪龍咬合力,高於之前的生物力學研究結果,介於4,100到8,200牛頓,這個數值大於許多現代肉食性哺乳動物,接近類似體型的鱷魚。
這群科學家也發現,恐爪龍的齒痕非常少見,而大型獸腳類恐龍的齒痕則較為常見,例如暴龍。所以他們推論,恐爪龍很少直接咬穿骨頭。恐爪龍可能只有在防禦或是獵捕獵物時,才會使用到它們的高咬合力[43]。
四肢功能

恐爪龍的最著名特徵是鐮刀狀的第二趾爪,但不同標本的第二趾爪的形狀與彎曲度都有所不同。奧斯特倫姆在1969年研究的正模標本,其第二趾爪是非常彎曲的,而在1976年研究的標本則較直,且與其他的趾爪很相似[44]。奧斯特倫姆認為這隻鐮刀狀趾爪的大小及形狀,隨者個體、性別或年紀的不同,而有所不同。目前已發現馳龍科在行走時,第二腳趾後縮、離地的解剖學[3]與足跡化石證據[45]。
奧斯特倫姆猜測恐爪龍是以前肢捉住獵物,並以鐮刀狀趾爪來割破肚子[19]。某些人員更提出,恐爪龍的鐮刀狀趾爪可劃開大型角龍類的身體[46]。但是後來的研究提出,這些鐮刀爪並非用作割開獵物的,而是刺戳獵物的[47]。另有研究人員參考恐爪龍、伶盜龍的腳部結構,使用機械模型攻擊豬屍體,但只能在屍體刺出淺穴,無法有效劃開肉體。研究人員假設,鎌刀狀趾爪也可能用來爬上動物的身體[48]。
在1970年,約翰·奧斯特倫姆曾參考鴕鳥與食火雞,發現這些鳥類會用較大型的第二趾爪攻擊獵物,進而產生恐爪龍會利用第二趾爪攻擊獵物的理論[19]。食火雞的第二趾爪可長達12.5公分[49]。奧斯特倫姆並引用一個1958年的鳥類論文,認為後肢與第二趾爪,可用來將獵物的身體撕開、將內臟挖出[50]。在近年,Chhristopher P. Kofron研究241件食火雞的攻擊紀錄,發現總計有1個人類、2隻狗的被攻擊死亡案例,但沒有證據顯示食火雞會用第二趾爪將獵物的內臟挖出[51][52]。相反地,食火雞將趾爪作為防衛武器,攻擊對牠們產生威脅的動物、或威嚇對方[49]叫鶴也具有較大的第二趾爪,卻是用在撕裂小型獵物,以適合吞嚥。[53]。在2011年的一項恐爪龍類趾爪研究,也認為第二趾爪無法作出有效的刺穿、劃開獵物身體的動作,並提出第二趾爪有固定獵物功能的理論[41]。

在2002年,肯尼思·卡彭特(Kenneth Carpenter)的生物力學研究,確認了恐爪龍的前肢最可能是用來捉住獵物的,因為其前肢的長度較其他獸腳亞目更易於捕捉。大及長的鳥喙骨顯示,恐爪龍有強壯的前肢肌肉,更加強了這個解釋[54]。卡彭特的研究也製作骨頭模型來實驗。實驗證明恐爪龍是不能掘曲牠的手臂,像鳥類般緊貼身體,這與嘉克斯·高斯特(Jacques Gauthier)[55]及葛瑞格利·保羅(Gregory S. Paul)兩人在80年代的各別推論有所不同[4]。
在2006年,菲力·森特(Phil Senter)指出恐爪龍的前肢未必只用來捕捉獵物,也可以將物體抓向胸部前方。恐爪龍被認為極可能前肢具有羽毛,但這會妨礙前肢的活動範圍。例如,當恐爪龍將前肢往前伸直時,掌心會自動翻轉朝前。當兩個前肢同時伸直時,前肢的羽毛會妨礙行動。所以森特認為,當恐爪龍在同時間只能用一隻手抓近物體,而不能同時用雙手。羽毛可能也限制了手指的功能,所以可能只有第三指可發生作用,例如在探挖土中的小型獵物時,也只有第三指可做出90度的彎曲[56]。在2001年,Alan Gishlick也對恐爪龍的前肢力學做了研究,他認為即使前肢具有羽毛,它們並不會明顯影響前肢的抓取能力。而且,當手掌抓取時,可與前肢作90度垂直,並可做類似擁抱的動作。這些發現與卡彭特與森特的前肢研究一致[57]。
在2006年,克里斯登·帕森斯(Kristen Parsons)提出恐爪龍的幼年及接近成年標本,與成年的標本有著一些形態上的不同。例如,幼年個體的手臂在比例上較成年的為長,顯示可能幼龍與成年的行為模式可能也不同[58]。另一個證據是腳趾趾爪的功能。帕森斯提出幼年個體的趾爪比較彎曲,可能有助於翻爬樹幹;成年個體的趾爪比較筆直,有助於在地面移動[59]。這個理論是根植於小型馳龍科可能會使用趾爪在攀爬樹幹[48]。
速度
馳龍科,尤其是恐爪龍,一直都被大眾媒體認為是奔走快速的恐龍,而奧斯特倫姆亦在原先的描述中有此推論[19]。但是在1969年首次描述時,並沒有發現完整的恐爪龍後腳,而奧斯特倫姆所估計的股骨長度,後來被證實是過長。奧斯特倫姆在1976年的研究注意到,在評估速度上,腳掌與脛骨的相對長度,比股骨與脛骨的比例更為重要。現今奔走快速的鳥類,如鴕鳥,腳掌與脛骨比例為0.95;而在恐龍中,似鴕龍是0.68,恐爪龍則是出奇的低,只有0.48。他於是結論出恐爪龍相對於其他恐龍並不怎麼快速,且更不是現今不能飛的鳥類的對手[44]。
恐爪龍的腳掌/脛骨比例低,部份是因牠非常短的蹠骨。小型個體的腳掌/脛骨比例,仍比較大個體的為長。奧斯特倫姆認為短的蹠骨是與鐮刀爪的功能有關,當年紀漸長時蹠骨的比例會更為短小,更支持他的理論。他解釋所有這些特徵(短的二趾上有大的爪、短的蹠骨等)都是為使後肢成為攻擊性的武器,鐮刀爪可以向上及向下攻擊,而腳則同時向後及向下拉,把獵物割破及撕開。奧斯特倫姆指短的蹠骨減少了腳骨在攻擊時的整體壓力,而恐爪龍的特殊肌肉構造是用作獵食而非奔跑或行走。所以他斷定恐爪龍的腳是平衡奔跑,及減低作為武器時遭受的壓力[44]。
在1981年,Richard Kool對於加拿大恐龍足跡化石的研究中,他針對英屬哥倫比亞蓋森組所發現的不同物種的足跡,做了初略的行走速度估計值。其中一個可能是恐爪龍留下的足跡,被估計以時速10.1公里行走[60]。
蛋化石

在2000年,從正模標本周遭石塊鑑定出來的恐爪龍蛋,可讓研究人員與其他獸腳亞目的蛋結構、巢穴及繁殖作出比較。在2006年,Tinner與馬克維奇評估這些蛋殼標本的時候,他們提出不同的假設,例如馳龍科是吃蛋的,或者蛋碎片與該隻恐爪龍是碰巧在一起的。因為這些蛋殼是在恐爪龍的腹肋與前肢之間,並非是在胃部之內,故這些蛋並非牠們的食物。而根據蛋殼的擠壓與破碎狀況,可推論蛋是完整埋入土中的,而在化石化的過程中破裂。另外,由於蛋周圍的骨骼並非分散或分開的,反而是與在生時差不多的姿勢,可見這個位點在化石化期間並沒有受到影響,故巧合之說亦不成立。事實上,腹肋通常很難發現保持天然狀態的,可見這些蛋是在埋葬時就在恐爪龍的腹下。這可能是恐爪龍的孵蛋或築巢行為,像傷齒龍科或竊蛋龍科;或是該恐爪龍死時,蛋仍在輸卵管之中[17]。
根據這些蛋的微結構,發現它們是屬於獸腳亞目,因為它們有著其他獸腳亞目蛋的特徵,並與鳥臀目及蜥腳下目蛋的不同。與其他手盜龍類比較,這些蛋化石較像竊蛋龍科的蛋,而非傷齒龍科的蛋。雖然該蛋經已碎裂,而很難評估其大小,但Tinner與馬克維奇根據蛋所經過的骨盆腔,推測出蛋的直徑約為7厘米。這個大小與最大的葬火龍蛋的直徑7.2厘米接近。而葬火龍與恐爪龍的體型接近,支持了蛋的估計值。再者,葬火龍蛋殼及恐爪龍蛋殼厚度差不多完全一致,由於蛋殼厚度影響著蛋的體積,故此有指牠們蛋的大小是完全一樣的[17]。
病理
在2001年,布魯斯·羅斯柴爾德(Bruce Rothschild)等人發表一份獸腳類恐龍的壓力性骨折研究。他們研究恐爪龍的43個手掌骨頭、52個腳掌骨頭,沒有發現壓力性骨折的跡象[61]。
編號YPM 5205標本的第二腳趾第二趾骨,被發現有骨折後癒合的跡象[62]。
大众文化
自从恐爪龙被发现并描述之后,就一直频繁地作为驰龙科的代表物种出现在各种科普读物和电视电影当中。恐爪龙出现在探索频道的纪录片《恐龙战争》中,片中两只恐爪龙齐心协力杀死了亚成年的波塞东龙。现实中,恐爪龙与波塞东龙并没有证据表明它们之间存在捕食关系。在2008年的纪录片《侏罗纪格斗俱乐部》中,一群恐爪龙杀死一只肌腱龙。
至於電影《侏罗纪公园》系列中伶盜龍(Velociraptor)卻類似恐爪龙的表现,後來在續集《侏罗纪世界》剧中借負責恐龍基因的吳博士之口解釋,複製恐龍是迎合人們對恐龍印象製作出來的,並非原本樣貌。
在電玩遊戲《侏羅紀世界:進化》及其續作中皆有出現恐爪龍,頭上長有冠飾,尾巴上還有類似帆的構造。
其他
參考資料
- Ostrom, J. H. (1970) "Stratigraphy and paleontology of the Cloverly Formation (Lower Cretaceous) of the Bighorn Basin area, Wyoming and Montana", Bulletin of the Peabody Museum of Natural History 35:1–234.
- (英文)Norell, Mark A.; & Makovicky, Peter J. . American Museum Novitates. 1999, 3282: 1–45.
- Ostrom, J. H. . Bulletin of the Peabody Museum of Natural History. 1970, 35: 1–234.
- (英文)Paul, G.S. . New York: Simon and Schuster. 1988: 366-369. ISBN 0-671-61946-2.
- (英文)Maxwell, W. D. & Witmer, L. M. . Journal of Vertebrate Paleontology. 1996, 16 (3): 51A.
- (英文)Witmer, Lawrence M. & Maxwell, William D. . Journal of Vertebrate Paleontology. 1996, 16 (3): 73A.
- (英文)Norell MA, Makovicky PJ. . Weishampel DB, Osmólska H, Dodson P (编). . University of California Press. 2004: 196–210. ISBN 978-0-520-24209-8.
- (英文)Norell, M.A.; Clark, J.M., Turner, A.H., Makovicky, P.J., Barsbold, R., and Rowe, T. . American Museum Novitates. 2006, 3545: 1–51.
- (英文)Turner, A.S.; Hwang, S.H.; and Norell, M.A. (PDF). American Museum Novitates. 2007, 3557: 1–27 [2007-06-18]. (原始内容存档 (PDF)于2009-03-26).
- (英文)Barsbold, R. . The Joint Soviet-Mongolian Palaeontological Expedition, Transactions. 1983, 19: 1–117.
- (英文)Benton, Michael J. . Blackwell Publishing. 2004: 472 [2008-01-10]. ISBN 9780632056378. (原始内容存档于2008-10-19).
- (英文)Ostrom, J. H. . Bulletin of the Peabody Museum of Natural History. 1970, 35: 1–234.
- (英文)Brinkman, D. L.; R. L. Cifelli, and N. J. Czaplewski. . Oklahoma Geological Survey Bulletin. 1998, 146: 1–27.
- (英文)Chen, Z.-Q.; Lubin, S. . The Mountain Geologist. 1997, 34: 51–62.
- (英文)Burton, D.; Greenhalgh, B.W.; Britt, B.B.; Kowallis, B.J.; Elliott, W.S.; Barrick, R. . Geological Society of America Abstracts with Programs. 2006, 38 (7): 52.
- (英文)Lipka, T. R. . Lucas, S.G., Kirkland, J.I. and Estep, J.W. (编). . New Mexico Museum of Natural History and Science Bulletin, 14. Albuquerque: New Mexico Museum of Natural History and Science. 1998: 229–234. OCLC 40283894.
- (英文)Grellet-Tinner, G.; and Makovicky, P. . Canadian Journal of Earth Sciences. 2006, 43: 705–719.
- (英文)Norell, M. A.; Gaffney, E. S.; and Dingus, L. . New York: Knopf. 1995: 126-130. ISBN 0-679-43386-4.
- (英文)Ostrom, J. H. . Peabody Museum of Natural History Bulletin. 1969, 30: 1–165.
- Ostrom, J.H., 1969, "A new theropod dinosaur from the Lower Cretaceous of Montana", Postilla 128: 1-17
- (英文)Ostrom, John H. . Postilla, Peabody Museum of Natural History Bulletin. 1974, 165: 1–11.
- (英文)Ostrom, J.H. . Breviora. 1976, 439: 1–21 [2012-11-13]. (原始内容存档于2016-12-19).
- (英文). American Museum of Natural History. 2007 [2007-07-13]. (原始内容存档于2007-08-12).
- (英文)Makovicky, P. J.; Grellet-Tinner, G. . In Bravo, A.M. and T. Reyes (eds.) 1st international symposium on dinosaur eggs and babies,Isona i Conca Dellà Catalonia, Spain, 23–26 September 1999. 2000: 123–128.
- (英文)Grellet-Tinner, Gerard. . Papeis Avulsos de Zoologia. 2006, 46 (1): 1–10. doi:10.1590/S0031-10492006000100001.
- (英文)Erickson, Gregory M.; Curry Rogers, Kristina; Varricchio, David J.; Norell, Mark A.; and Xu, Xing. (pdf). Biology Letters. 2007, 3 (5): 558–61 [2007-07-26]. PMC 2396186
 . PMID 17638674. doi:10.1098/rsbl.2007.0254.
. PMID 17638674. doi:10.1098/rsbl.2007.0254. - (英文)Fastovsky DE, Weishampel DB. . Fastovsky DE, Weishampel DB (编). . Cambridge University Press. 2005: 265–299. ISBN 978-0-521-81172-9.
- (英文)Ostrom, J. H. . Biological Journal of the Linnean Society. 1976, 8: 91–182.
- (英文)Xu, X.; Zhou, Z.; Wang, X.; Kuang, X.; Zhang, F. & Du, X. . Nature. 2003, 421 (6921): 335–340. PMID 12540892. doi:10.1038/nature01342.
- (英文)Hwang, S. H.; Norell, M. A.; Ji, Q. & Gao, K. (PDF). American Museum Novitates. 2002, 3381: 44 [2022-08-03]. (原始内容存档 (PDF)于2022-02-26).
- (英文)徐星、汪筱林、吳肖春. . Nature. 16 September 1999, 401: 262–266 [2007-06-18]. (原始内容存档于2006-07-20).
- (英文)Turner, A.H.; Makovicky, P.J.; and Norell, M.A. (pdf). Science. 2007, 317 (5845): 1721 [2008-01-22]. (原始内容存档 (PDF)于2009-09-19).
- (英文)Prum, R.; Brush, A. H. . The Quarterly Review of Biology. 2002, 77: 261–295.
- Senter, P. . Journal of Vertebrate Paleontology. 2006, 26 (4): 897–906. doi:10.1671/0272-4634(2006)26[897:COFFBD]2.0.CO;2.
- (英文)Forster, C. A. . The Mosasaur. 1984, 2: 151–163.
- (英文)Wedel, M. J.; Cifelli, R. L. . Oklahoma Geology Notes. 2005, 65: 40–57.
- (英文)Maxwell, W. D.; Ostrom, J.H. . Journal of Vertebrate Paleontology. 1995, 15 (4): 707–712. doi:10.1080/02724634.1995.10011256.
- (英文)Brinkman, Daniel L.; Cifelli, Richard L., Czaplewski, Nicholas J. . Oklahoma Geological Survey. 1998, (164): 27.
- (英文)Roach, B. T.; D. L. Brinkman. . Bulletin of the Peabody Museum of Natural History. 2007, 48 (1): 103–138.
- Li, Rihui; Lockley, Martin G.; Makovicky, Peter J.; Matsukawa, Masaki; Norell, Mark A.; Harris, Jerald D.; Liu, Mingwei. . Naturwissenschaften. 2007, 95 (3): 185. doi:10.1007/s00114-007-0310-7.
- (英文)Fowler, D.W.; Freedman, E.A.; Scannella, J.B.; Kambic, R.E. . PLoS ONE. 2011, 6 (12): e28964 [2012-11-15]. PMID 22194962. doi:10.1371/journal.pone.0028964. (原始内容存档于2015-01-02).
- (英文)Therrien, F., Henderson, D.M. and Huff, C.B. (2005). "Bite me: biomechanical models of theropod mandibles and implications for feeding behavior." Pp. 179–237 in Carpenter, K. (ed.), The Carnivorous Dinosaurs. Indianapolis: Indiana University Press.
- (英文)Gignac, P.M.; Makovicky, P.J.; Erickson, G.M.; Walsh, R.P. . Journal of Vertebrate Paleontology. 2010, 30 (4): 1169–1177. doi:10.1080/02724634.2010.483535.
- (英文)Ostrom, J. H. . Breviora. 1976, 439: 1–21 [2012-11-13]. (原始内容存档于2020-09-27).
- (英文)Li, Rihui; Lockley, Martin G.; Makovicky, Peter J.; Matsukawa, Masaki; Norell, Mark A.; Harris, Jerald D.; Liu, Mingwei. . Naturwissenschaften. 2007, 95 (3): 185–91. PMID 17952398. doi:10.1007/s00114-007-0310-7.
- (英文)Adams, Dawn (1987) "The bigger they are, the harder they fall: Implications of ischial curvature in ceratopsian dinosaurs" pg 1–6 in Currie, Philip J. and Koster, E. (eds) Fourth symposium on mesozoic terrestrial ecosystems. Tyrrell Museum, Drumheller, Canada
- (英文)Carpenter, K. . Gaia. 1998, 15: 135–144.
- Manning, Phil L.; Payne, David; Pennicott, John; Barrett, Paul M.; Ennos, Roland A. . Biology Letters. 2006, 2 (1): 110–112. PMC 1617199 . PMID 17148340. doi:10.1098/rsbl.2005.0395.
- (英文)Davies, S. J. J. F. . New York: Oxford University Press. 2002. ISBN 0-19-854996-2.
- (英文)Gilliard, E. T. . Garden City, NY: Doubleday. 1958.
- (英文)Kofron, Christopher P. . Journal of Zoology. 1999, 249 (4): 375–381. doi:10.1111/j.1469-7998.1999.tb01206.x.
- (英文)Kofron, Christopher P. . Memoirs of the Queensland Museum. 2003, 49 (1): 335–338.
- (英文)Redford, Kent H.; Peters, Gustav. . Journal of Field Ornithology. 1986, 57 (4): 261–269. JSTOR 4513154.
- (英文)Carpenter, K. . Senckenbergiana Lethaea. 2002, 82: 59–76. doi:10.1007/BF03043773.
- (英文)Gauthier, J.; Padian, K. . M.K. Hecht, J.H. Ostrom, G. Viohl, & P. Wellnhofer (eds) (编). . The Beginnings Of Birds. Eischtatt: Freunde des Jura-Museums Eichstätt: 185–197. 1985. isbn 3-9801178-0-4.
- (英文)Senter, P. . Journal of Vertebrate Paleontology. 2006, 26 (4): 897–906. doi:10.1671/0272-4634(2006)26[897:COFFBD]2.0.CO;2.
- (英文)Gishlick, A.D. . Gauthier, J. and Gall, L.F. (编). . New Haven: Yale Peabody Museum. 2001: 301–318.
- (英文)Parsons, W.; Parsons, K. . Journal of Vertebrate Paleontology. 2006, 26 (3): 109A [2007-06-18]. (原始内容存档于2007-08-03).
- (英文)Parsons, W. L.; and Parsons, K. M. (PDF). Bulletin of the Buffalo Society of Natural Sciences. 2009, 38: 43–54 [2010-11-14]. (原始内容 (PDF)存档于2010-07-03).
- (英文)Kool, R. . Canadian Journal of Earth Science. 1981, 18 (4): 823–825. doi:10.1139/e81-077.
- Rothschild, B., Tanke, D. H., and Ford, T. L., 2001, Theropod stress fractures and tendon avulsions as a clue to activity: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, p. 331-336.
- (英文)Molnar, R. E., 2001, Theropod paleopathology: a literature survey: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, p. 337-363.