拟菌病毒属
拟菌病毒属(學名:)是一个包括Acanthamoeba polyphaga mimivirus(APMV)的一个种类,或许是与演化史相关的巨型病毒。[1]通常所说的“拟菌病毒”就指APMV。在口語中,APMV更普遍簡稱為“米米病毒”(mimivirus)。2011年10月中旬,科學家發現了巨大病毒,這是已知衣壳直径最大的病毒。[2]拟菌病毒比起其他病毒有着庞大的基因组。拟菌病毒(mimicking microbe)的命名反映了它的体积及革兰氏染色的特性。[3]
| 拟菌病毒属 | |
|---|---|
 | |
| 病毒分類 | |
| (未分级): | 病毒 Virus |
| 域: | 多變DNA病毒域 Varidnaviria |
| 界: | 班福病毒界 Bamfordvirae |
| 门: | 核質病毒門 Nucleocytoviricota |
| 纲: | 巨病毒纲 Megaviricetes |
| 目: | 模拟病毒目 Imitervirales |
| 科: | 擬菌病毒科 Mimiviridae |
| 属: | 拟菌病毒属 Mimivirus |
| 种 | |
| |
发现
拟菌病毒最初是在1992年的一次对军团病研究中,偶然在变形虫多食棘變形蟲(Acanthamoeba polyphaga)中发现的。然而在之后的革兰氏染色实验中被错误地认为是一种革兰氏阳性菌,并被命名为“布拉德福德球菌”(Bradfordcoccus)。到2003年,法国马赛地中海大学在《科学》期刊上发布了文章,认定这个有机体为一种病毒。[4]
分类
国际病毒分类委员会将拟菌病毒归为拟菌病毒科。[5]并在生物病毒分类中归为I类病毒(双链DNA病毒)。[6]在不严格的分类下,拟菌病毒加入核质巨DNA病毒(NCLDV)一类。这类病毒都有着类似的分子特性和庞大的基因组。拟菌病毒同样有21个同源于NCLDV的高度保守基因,并且进一步的研究也显示拟菌病毒可能是NCLDV的一个分支。[4]
结构
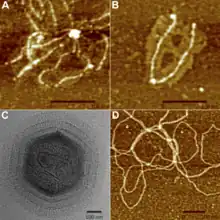
擬菌病毒的衣壳直徑達400nm,是已知第三大的病毒,僅次於潘多拉病毒和近期发现的巨大病毒。加上衣壳表面的蛋白丝,总长度可达600nm. 各种文献对此的描述高度相似,病毒大小都在400nm到800nm之间。在电子显微镜下,病毒的衣壳呈六边形,因此衣壳的立体结构为正二十面體。[7]拟菌病毒并没有包膜,这意味着这类病毒并非通过胞吐被放出细胞的。[8]
发现拟菌病毒的小组后来发现了一种略微更大的病毒,命名为妈妈病毒(mamavirus) ,以及能感染它的亚病毒Sputnik virophage.[9]
拟菌病毒有着所有NCLDV上共同的形态学特征。M. Suzan-Monti等人提出,如同其他NCLDV病毒,拟菌病毒可能也有这内含的包裹DNA核的脂质层。在电子显微镜下,病毒体浓缩的核呈现出黑色。病毒庞大的基因组就在这个区域之中。
一些mRNA的转录子可以被拟菌病毒独立完成。如同其他NCLDV的DNA聚合酶的转录,一种衣壳蛋白质和一个类似于TFII的转录因子也被发现。然而,三种明显不同的氨酰-tRNA合成酶的转录子以及4种未知的拟菌病毒独有的mRNA分子也被发现。这些预先打包的转录子能够在病毒基因没有表达的情况下被翻译,并且可能对拟菌病毒的复制是必要的。其他的DNA病毒,如人类巨细胞病毒和单纯疱疹病毒I型也有mRNA预先打包转录的特点。[8]
基因组
拟菌病毒的基因组是一个线性的,有着1,181,404对碱基的DNA双链。[10]是目前已知病毒中基因组最庞大的一种,超出了仅次于它的伦比约餐厅虫病毒(Cafeteria roenbergensis virus)450,000对碱基。此外,它甚至比至少30种细胞体还大。[11]
除了庞大的基因组,拟菌病毒还有着大约979个蛋白质编码基因,远远超出了病毒最低所需的4个基因(参见 MS2噬菌体和Qβ噬菌体)。[12]分析显示,拟菌病毒特有的基因包括氨酰tRNA合成酶以及其他一些早前被认为只能由细胞微生物编码的酶。正如其他的巨型DNA病毒,拟菌病毒也被发现有着其他病毒不具有的与糖类、脂类和氨基酸代谢相关的基因。[8]大约90%的基因组是编码基因,而另外10%为非编码DNA。
复制
拟菌病毒的复制过程尚不清楚。但是我们至少知道此病毒能与阿米巴细胞膜上的一种受体结合。一旦进入阿米巴,便进入一段隐蚀期,这时病毒会消失,细胞会正常工作。在感染约4h后,细胞表面会显现出一些聚集物。感染8h后,在细胞中的拟菌病毒便清晰可见。细胞质会持续被新组装的病毒体填充。感染约24h后,细胞便极为可能破裂,并释放出新的拟菌病毒病毒体。[8]
目前我们对拟菌病毒的复制周期几乎一无所知,显然是附着于细胞表面,进入,病毒核物质释放,DNA复制,转录,翻译,组装,最后释放子病毒体。然而,研究人员是使用对被感染的细胞的显微照相来得出上述综述的。这些显微照片显示了拟菌病毒的衣壳在细胞核中组装,并在出核时获得一个脂质膜,以及一些与其他包括NCLDV的不少病毒相似的颗粒。这些颗粒在其他病毒中被称为病毒工厂(viral factories),它能使宿主细胞被更多地改造,并病毒的组装更快。
致病性
拟菌病毒或许是某些肺炎的病原体。这种推测是基于在某些肺炎患者体内发现此病毒抗体的间接证据。[13]然而,将拟菌病毒归为病原体的理论是极为脆弱的,因为仅有极少的几篇文章报告了拟菌病毒与肺炎的潜在关系。造成肺炎的最大因素目前还尚未知晓。[14]即便如此,已经有一种拟菌病毒从一名突尼斯的女性肺炎患者中分离出来。[15]
对生命定义的引思
拟菌病毒的许多特性将之放在了生命与非生命之间。它并不比某些细菌小,如康氏立克次氏体(Rickettsia conorii)和Tropheryma whipplei,并且有着与不少细菌相当甚至更多的基因。在之前病毒还没有被认为能够进行编码。此外,拟菌病毒还有着核苷酸和氨基酸合成相关的基因,这甚至是一些小型的专性寄生细菌所不具有的。然而,拟菌病毒确实缺少了核糖體蛋白質,这使得他们必须依靠宿主细胞的蛋白转运和能量代谢。
因为拟菌病毒的起源非常古老,甚至有可能先于细胞[16][17],拟菌病毒已经在生命起源的争论中成为一种论据。一些拟菌病毒独有的基因,包括那些衣壳相关的基因,在多种能感染各域生物的病毒中保留。这暗示了拟菌病毒与一种先于细胞生物并且在地球生物的起源中起重要作用的DNA病毒有联系。[16]另一种假设是,有明显的三种DNA病毒与目前的三个域(真核生物、古生菌、细菌)的产生有关。[17]也有猜想认为这些病毒可能一开始是细菌,后来退化成了病毒。
参见
| 維基物種上的相關:拟菌病毒属 |
- 伦比约餐厅虫病毒Cafeteria roenbergensis virus (CroV)最大的海洋病毒
- 马赛病毒Marseillevirus—另一种巨型病毒
- 百萬病毒Megavirus—另一种巨型病毒
- 生殖支原体Mycoplasma genitalium—已知最小的細菌之一
- 纳古菌Nanoarchaeum equitans—最小的独立生存的细胞
- 纳米细菌Nanobacterium
- 纳诺比Nanobe
- 非细胞生物
- 潘多拉病毒Pandoravirus—已知的最大病毒
- 细小病毒Parvovirus—已知的最小的病毒
- 遍在遠洋桿菌Pelagibacter ubique—拥有最小的基因组的细菌
- 噬病毒体Virophage—一种需要宿主细胞一同感染巨型细胞的病毒
参考文献
- Elodie Ghedin, Jean-Michel Claverie. . Virology Journal. 2005-08-16, 2: 62 [2018-04-02]. ISSN 1743-422X. doi:10.1186/1743-422x-2-62.
- . London: Telegraph UK. 11 October 2011 [11 November 2011]. (原始内容存档于2014-01-15).
- Wessner, D R. . Nature Education. 2010, 3 (9): 61 [2012-01-07]. (原始内容存档于2020-11-01).
- La Scola B, Audic S, Robert C, Jungang L, de Lamballerie X, Drancourt M, Birtles R, Claverie JM, Raoult D. . Science. 2003, 299 (5615): 2033. PMID 12663918. doi:10.1126/science.1081867.
- Claverie J-M. Mahy W.J. and Van Regenmortel M. H. V. , 编. 1. Oxford: Academic Press. 2010: 189.
- Leppard, Keith; Nigel Dimmock; Easton, Andrew. 6. Blackwell Publishing Limited. 2007: 469–470.
- Xiao C, Kuznetsov YG, Sun S, Hafenstein SL, Kostyuchenko VA, Chipman PR, Suzan-Monti M, Raoult D, McPherson A, Rossmann MG. Sugden, Bill , 编. . PLoS Biol. 2009-04-28, 7 (4): e92. PMC 2671561
 . PMID 19402750. doi:10.1371/journal.pbio.1000092.
. PMID 19402750. doi:10.1371/journal.pbio.1000092. - Suzan-Monti M, La Scola B, Raoult D. . Virus Research. 2006, 117 (1): 145–155. PMID 16181700. doi:10.1016/j.virusres.2005.07.011.
- Nature: "'Virophage' suggests viruses are alive" (页面存档备份,存于)by Helen Pearson 2008
- . NCBI.
- Claverie, Jean-Michel; et al.. . Virus Research. 2006, 117 (1): 133–144. PMID 16469402. doi:10.1016/j.virusres.2006.01.008.
- Prescott, Lansing M. 2nd edition , 编. . Dubuque, IA: Wm. C. Brown Publishers. 1993. ISBN 0-697-01372-3.
- La Scola B, Marrie T, Auffray J, Raoult D. . Emerg Infect Dis. 2005, 11 (3): 449–52 [2013-12-10]. PMC 3298252 . PMID 15757563. doi:10.3201/eid1103.040538. (原始内容存档于2008-10-19).
- Marrie TJ, Durant H, Yates L. . Reviews of Infectious Diseases. 1989, 11 (4): 586–99. PMID 2772465. doi:10.1093/clinids/11.4.586.
- Saadi, H; Pagnier, I; Colson, P; Cherif, JK; Beji, M; Boughalmi, M; Azza, S; Armstrong, N; Robert, C; Fournous, G; La Scola, B; Raoult, D. . Clinical infectious diseases : an official publication of the Infectious Diseases Society of America. 2013-06-21, 57 (4): e127–34. PMID 23709652. doi:10.1093/cid/cit354.
- Siebert, Charles. . Discover Magazine. 2006-03-15. (原始内容存档于2019-10-26).
- Forterre, Patrick. . PNAS. 2006, 103 (10): 3669–3674. PMC 1450140 . PMID 16505372. doi:10.1073/pnas.0510333103.
补充资料
- Raoult, D.; et al.. . Science. 2004, 306 (5700): 1344–1350. PMID 15486256. doi:10.1126/science.1101485.
- Ghedin, Elodie; Claverie, J. M. . Virology Journal. 2005, 2: 62. PMC 1215527 . PMID 16105173. doi:10.1186/1743-422X-2-62.
- Peplow, Mark, 2004, "Giant virus qualifies as 'living organism' (页面存档备份,存于)," News@Nature
- . Press Release. Paris: Centre national de la recherche scientifique. 2003-03-28. (原始内容存档于2004-06-03).
- New Scientist, Issue 2544, 25 March 2006.
- GiantVirus.org (页面存档备份,存于)
- Highfield, Roger. . London: Daily Telegraph. 2004-10-15 [2021-12-18]. (原始内容存档于2008-01-27).
- . Science News. 2009-04-28 [2013-12-10]. (原始内容存档于2012-01-07).
- Keim, Brandon. . Wired Science. 2009-05-05 [2013-12-10]. (原始内容存档于2012-11-02).
外部链接
- Viralzone: Mimiviridae (页面存档备份,存于)
- International Committee on Taxonomy of Viruses (ICTV) picture gallery (页面存档备份,存于)—images of mimivirus
- Giant Viruses: The recent discovery of really, really big viruses is changing views about the nature of viruses and the history of life (页面存档备份,存于) by James L. Van Etten July–August 2011 American Scientist Volume 99, Number 4, page: 304, doi:10.1511/2011.91.304
- The webpage for Mimivirus