大夏巨龍屬
大夏巨龍屬(屬名:,意為「大夏河的巨人」)是多孔椎龍類蜥腳恐龍的一個屬,屬於基礎泰坦巨龍類或其中的盤足龍科,化石來自中國西北部甘肅省蘭州盆地的下白堊紀地層,迄今已發現部分脊椎、肩帶和大腿骨[1]。其全長估計可達23至30公尺,在亞洲恐龍中名列前茅[1][2];而體重估計達23噸,和許多同等長度級的近親相比則顯小[3]。大夏巨龍具有極長的頸部,頸椎的大小在基礎泰坦巨龍類中也是數一數二,重建後的頸部可能超過12公尺,幾乎佔去全長的一半[1][3]。
| 大夏巨龙属 化石时期:白堊紀早期, | |
|---|---|
 | |
| 臺灣國立自然科學博物館展出的骨架模型 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 亚目: | †蜥脚形亚目 Sauropodomorpha |
| 下目: | †蜥腳下目 Sauropoda |
| 演化支: | †巨龍形類 Titanosauriformes |
| 演化支: | †多孔椎龙类 Somphospondyli |
| 科: | †盘足龙科 Euhelopodidae |
| 属: | †大夏巨龙属 Daxiatitan You et al., 2008 |
| 模式種 | |
| †炳靈大夏巨龍 Daxiatitan binglingi You et al., 2008 | |
發現及命名

大夏巨龍是繼劉家峽黃河巨龍之後,第二個在蘭州盆地河口群發現的蜥腳類。模式種炳靈大夏巨龍(Daxiatitan binglingi)是在2008年由尤海魯、李大慶、周伶琦、季強所敘述、命名。屬名意指其發現地,即流經甘肅臨夏的大夏河,加上希臘神話的泰坦組成;種名炳靈來自劉家峽大壩附近著名的炳靈寺。[1]
正模標本(編號GSLTZP03-001)是一個部分骨骼,包含後10節頸椎、10節背椎、2節近端尾椎、部分頸肋和背肋、1個脈弓、右肩胛骨、右鳥喙骨、右股骨;現保存於蘭州的甘肅省地礦局第三地質礦產勘查院古生物研究開發中心。[1]
敘述
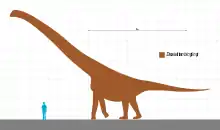

盤足龍是估計大夏巨龍薦前脊椎數目的良好參考範例,因為同樣位於泰坦巨龍類演化的基礎位置且是唯一有保存完整薦前脊椎(含薦椎)的物種。盤足龍具有31節薦前脊椎(19節頸椎加12節背椎)以及6節薦椎,而大夏巨龍與盤足龍具有類似的頸椎長度變化模式,因此可推論大夏巨龍的頸部也像盤足龍那樣的加長;大夏巨龍最後九節頸椎長度為6.1公尺,同於盤足龍的比例,於是估計大夏巨龍頸部全長12.2公尺。根據同樣加長頸部的馬門溪龍頸部佔全長比例為47%來推估,大夏巨龍身長約為26公尺,而重建之後加上脊椎間距的全長可達到30公尺,這將使大夏巨龍成為中國已知最長的恐龍之一;同時也可能是最重的。大夏巨龍的頸椎椎體關節面大致呈半球形,且關節面/椎體長的比值高,最長的頸椎關節面寬和高分別為28公分及31公分,而椎體長91公分,代表大夏巨龍的頸部圓潤而肥碩,相比之下馬門溪龍具有較低的值(依序為14、30、73公分),頸部較狹窄纖細,因此可推測大夏巨龍體重可能高於體長相近的馬門溪龍。[1]
大夏巨龍具有以下鑑定特徵(自衍徵),不同於其他巨龍形類:頸椎的前關節面後移;肩胛骨骨體的遠緣與其長軸斜交;股骨遠端髁相對於股骨體向背側方傾斜10度,且從遠端來看該遠端髁處於後內—前側方向。另外大夏巨龍具有獨特的股骨構造使步行時後肢呈現強烈外撇的姿態,這能夠從劉家峽地區同一層位發現的強烈外轉、寬間距的蜥腳類腳印得到佐證。[1]
分類
支序分類學研究分析將大夏巨龍置於泰坦巨龍類的基礎位置,比盤足龍衍化,但比馬拉威龍原始;因此大夏巨龍對於研究泰坦巨龍類的起源和早期演化具有重要意義。[1]以下演化樹擷取自Mannion等人(2019)對於白堊紀亞洲泰坦巨龍類的重新檢視:[4]
| 巨龍形類 Titanosauriformes |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
古生物學
李立國(2019)針對蜥腳類頸部的肌肉與肌腱系統與現代鳥類的頸部結構作比較,其中對大夏巨龍進行了重新檢視與詳盡的描述。[5]
雖然大夏巨龍的骨骼保存得不完整,但保存的脊柱關節仍相連,顯示這些部位生前曾有強韌的組織連接著;平行的胸肋、關節脫落的肢骨則代表這些部位在掩埋前受到古生物學上的干擾而分離。針對大夏巨龍的脊柱和附肢骨骼進行檢驗,提供了許多解剖學上的線索,來重建用作連接功能的軟組織。神經弓椎板複雜的「N」與「M」形態,加上背椎側面數道彎脊,可能曾是肌腱與軟組織附著的地方。椎體側面大量的氣孔表示具有頸部氣囊腔室,這些腔室被認為由三個氣囊組成的系統所填充。[5]
_white_form.JPG.webp)

研究根據一些現代鳥類有著很大不同的骨學與肌理學形式,來進行比較並重建出大夏巨龍頸部腹側的肌腱系統。取材參考的鳥類包含大藍鷺:頸部肌肉長而細瘦(LL型/long light configuration)、小天鵝:頸部肌肉短而粗壯(SS型/short strong configuration)這兩種對立形式的頸部肌肉結構。蒼鷺LL型的重疊肌腱形式屬於長肌腱-細肌肉,在骨學上以第19節頸椎動脈突的深滑槽作為對應證據;與軀幹相對位移的頸部肌肉、神經、血管以生物力學方式作為「遠端神經中樞」來控制整個頸部,因此主要的長肌腱、軀幹中央的腹部肌肉、扭轉的S形頸部由單一肌群、頸長肌以及三節關鍵脊椎所控制。相反地,天鵝SS型的人字形式屬於短肌腱-粗肌肉,加上頸動脈突缺乏凹槽,代表頸部具有柔韌性,可以分別控制單一節頸椎而非整個脖子;肌肉均勻的天鵝頸部也佔體重較多比例;在SS人字形式中,肌腱、血管、神經系統共同作用構成一個集合網絡,能夠個別控制頸椎以增加靈活度。由以上兩種形式的比較,顯示大夏巨龍和其他類似的蜥腳類極度加長的頸肋可能具有覓食或防禦功能。在運動性與體重平衡上,長頸部的蜥腳類,包含大夏巨龍、馬門溪龍等,較符合LL型的肌腱主導型式;而短頸肋的蜥腳類,包含梁龍、迷惑龍等,則較符合SS型的肌肉主導型式。[5]
古生態學

與大夏巨龍同樣出土於蘭州盆地河口群的黃河巨龍,兩者在肩胛骨形態上有足夠差異,因此各自為獨立的屬,表示當地至少同時存在著兩種以上的蜥腳類物種,如同其他常見蜥腳類的地層(如侏羅紀晚期北美的莫里遜組和中國的沙溪廟組)代表這種情況並不罕見,可能是因為多種蜥腳類各自佔據不同生態棲位而得以共存,至於大夏巨龍和黃河巨龍彼此間的棲位差異則尚待進一步考證。[1]
參考資料
- You, H.-L.; Li, D.-Q.; Zhou, L.-Q.; Ji, Q. . Gansu Geology. 2008, 17 (4): 1–10 [2020-10-03]. ISSN 1004-4116. (原始内容存档于2020-04-01).
- Holtz, T. R. Jr. (PDF). . Indiana University Press. 2012 [2018-07-04]. ISBN 0-375-82419-7. (原始内容存档 (PDF)于2017-08-12).
- Molina-Pérez, R.; Larramendi, A. . (譯) Donaghey, J. Princeton University Press. 2020: 123, 160, 262. ISBN 978-0-691-19069-3.
- Mannion, P.D.; Upchurch, P.; Jin, X.; Zheng, W. . Royal Society Open Science. 2019, 6 (8): 191057. Bibcode:2019RSOS....691057M. PMC 6731702
 . PMID 31598266. doi:10.1098/rsos.191057.
. PMID 31598266. doi:10.1098/rsos.191057. - Liguo, Li. . Drexel University: 21–23. September 2019.
- . [2021-03-26]. (原始内容存档于2020-02-04).