网状结构
网状结构(英語:)又称网状系统[2],是在中枢神经系统中,一系列介于脊髓上端到丘脑之间,由白质和灰质交织形成的弥散性神经网络。其涉及延髓中央、脑桥被盖、中脑、下丘脑等多个部位,大部分位于脑干内。
| 网状结构 | |
|---|---|
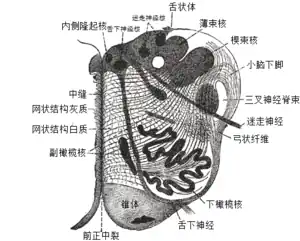
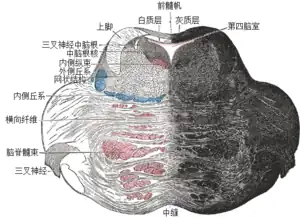 | |
 关于橄榄体中部的对延髓进行的截面(网状结构灰质与网状结构白质被标注在左侧) | |
| 基本 | |
| 位置 | 脑干、下丘脑等 |
| 标识字符 | |
| 拉丁文 | formatio reticularis |
| MeSH | D012154 |
| NeuroNames | 1223 |
| NeuroLex ID | nlx_143558 |
| TA98 | A14.1.00.021、A14.1.05.403、A14.1.06.327 |
| TA2 | 5367 |
| FMA | FMA:77719 |
| 格雷氏 | p.784 |
| 《神经解剖学术语》 | |
网状结构内除了有多个界限清楚且功能明确的神经细胞核团、神经纤维束外,另有散在分布的神经元群及纵横交错的神经纤维,它们共同形成一组复杂网络。网状结构内的神经细胞形状也很复杂,大小不等,轴突较长,侧枝较多。其功能涉及觉醒/睡眠循环,以及肌肉张力、心脏反射,并可以过滤进入的刺激以区分无关的背景刺激;因此网状结构对于高等生物控制一些身体基本功能是必须的,并且是脑部系统发生学上最老的部分之一。
网状结构的神经元,特别是上升网状激活系统的神经元,在维持行为唤醒和意识方面发挥着至关重要的作用。网状结构的总体功能是调节和前运动,涉及躯体运动控制、心血管控制、疼痛调节、睡眠和意识以及习惯化。调节功能主要存在于网状结构的喙部,而运动前功能则位于更多尾部区域的神经元中。
网状结构分为三列:中缝核(中缝核)、巨细胞网状核(内侧区)和小细胞网状核(外侧区)。中缝核是合成神经递质血清素的地方,在情绪调节中起着重要作用。巨细胞核参与运动协调。小细胞核调节呼气。
网状结构对于控制高等生物的一些基本功能至关重要,并且是系统发育上最古老的大脑部分之一。
脑干网状系统是一系列位於脑干內互相連結的神經核。脑干内除了有多个界限清楚且功能明确的神经细胞核团、神经纤维束外,其余散在分布的神经元群加上纵横交错的神经纤维,位于脑干核心的一组复杂网络,即为脑干网状结构。
结构

人类的网状结构由近 100 个脑核组成,并包含许多前脑、脑干和小脑等区域的投射。网状丘脑投射纤维、弥漫性丘脑皮质投射、上行胆碱能投射、下行非胆碱能投射和下行网状脊髓投射。网状结构还包含两个主要的神经子系统,即上升网状激活系统和下降网状脊髓束,它们介导不同的认知和生理过程。它在矢状面和冠状面均已被功能性劈裂。
传统上,网状核分为三列:
- 在中柱 - 中缝核
- 在中间柱 - 巨细胞核(因为细胞尺寸较大)
- 在侧柱中 - 细细胞核(因为细胞尺寸较小)
最初的功能分化是尾侧和喙侧的划分。这是基于这样的观察:吻侧网状结构的损伤会引起猫脑的睡眠过度。相反,网状结构尾部的病变会导致猫失眠。这项研究得出了尾部抑制网状结构的头端部分的想法。
矢状划分揭示了更多形态学差异。中缝核在网状结构的中间形成一个脊,并且直接到其外围,有一个称为内侧网状结构的部分。内侧射频较大,具有长的上升和下降纤维,并被外侧网状结构包围。外侧射频靠近脑神经的运动核,主要介导其功能。
功能
网状结构由 100 多个小型神经网络组成,具有多种功能,包括:
- 躯体运动控制——一些运动神经元将轴突发送到网状形成核,产生脊髓的网状脊髓束。这些束的作用是维持音调、平衡和姿势 – 尤其是在身体动作时。网状结构还将眼睛和耳朵的信号传递给小脑,以便小脑可以在运动协调中整合视觉、听觉和前庭刺激。其他运动核包括凝视中心(使眼睛能够跟踪和注视物体)和中央模式发生器(产生呼吸和吞咽的节律信号)。
- 心血管控制——网状结构包括延髓的心脏和血管运动中心。
- 疼痛调节——网状结构是下半身疼痛信号到达大脑皮层的一种方式。它也是下行镇痛通路的起源。这些通路中的神经纤维在脊髓中发挥作用,阻止某些疼痛信号向大脑的传递。
- 睡眠和意识——网状结构对丘脑和大脑皮层有投射,使其能够对哪些感觉信号到达大脑并引起我们的意识注意施加一定的控制。它在警觉和睡眠等意识状态中发挥着核心作用。网状结构损伤可导致不可逆的昏迷。
- 习惯化——这是大脑学会忽略重复的、无意义的刺激,同时对他人保持敏感的过程。一个很好的例子是,一个人可以在大城市的喧闹交通中睡觉,但会因警报声或婴儿哭声而立即被吵醒。调节大脑皮层活动的网状形成核是上行网状激活系统的一部分。
主要子系统
上行网状激活系统

上行网状激活系统(ARAS)也称网状激活系统(RAS),是脊椎动物大脑中一组相互连接的核团,负责调节觉醒和睡眠-觉醒转换。 ARAS是网状结构的一部分,主要由丘脑中的各种核团和一些多巴胺能、去甲肾上腺素能、血清素能、组胺能、胆碱能和谷氨酸能脑核团组成。
ARAS 的结构
ARAS 由多个神经回路组成,通过穿过丘脑和下丘脑的不同通路将后中脑和前脑桥的背侧部分连接到大脑皮层。 [3] [4] [5] ARAS 是不同核团的集合——上脑干、脑桥、延髓和下丘脑后部每侧都有 20 多个核团。这些神经元释放的神经递质包括多巴胺、去甲肾上腺素、血清素、组胺、乙酰胆碱和谷氨酸。 [3] [6] [4] [5]它们通过直接轴突投射和丘脑中继的间接投射来施加皮质影响。 [4] [5] [7]
丘脑通路主要由脑桥被盖中的胆碱能神经元组成,而下丘脑通路主要由释放单胺神经递质(即多巴胺、去甲肾上腺素、血清素和组胺)的神经元组成。 [3] [6] ARAS 中释放谷氨酸的神经元相对于单胺能和胆碱能神经元的发现要晚得多。 [8] ARAS 的谷氨酸能成分包括下丘脑的一个核和多个脑干核。 [4] [8] [9]下丘脑外侧的食欲素神经元支配上行网状激活系统的每个组成部分并协调整个系统内的活动。 [5] [10] [11]
| 细胞核类型 | 介导觉醒的相应核 | 来源 |
|---|---|---|
| 多巴胺能 细胞核 |
|
[3] [6] [4] [5] |
| 去甲肾上腺素能 细胞核 |
|
[3] [6] [5] |
| 血清素能 细胞核 |
|
[3] [6] [5] |
| 组胺能 细胞核 |
|
[3] [6] [12] |
| 胆碱能 细胞核 |
|
[3] [4] [5] [8] |
| 谷氨酸能 细胞核 |
|
[4] [5] [8] [9] [12] [13] |
| 丘脑 细胞核 |
|
[3] [4] [14] |
ARAS 由进化上古老的大脑区域组成,这些区域对于动物的生存至关重要,并在逆境期间受到保护,例如在托塞尔反射(又名“动物催眠”)的抑制时期。 [upper-alpha 1] [16]上行网状激活系统,将神经调节投射发送到皮层 - 主要连接到前额叶皮层。 [17]与皮质运动区域的连接似乎较低。 [17]
调节睡眠-觉醒转换
ARAS 的主要功能是修改和增强丘脑和皮质功能,从而导致脑电图(EEG) 不同步。 [upper-alpha 2] [20] [21]清醒和睡眠期间大脑的电活动存在明显差异:低电压快速突发脑电波(EEG 去同步)与清醒和快速眼动睡眠(电生理学上相似)相关);在非快速眼动睡眠期间会发现高压慢波。一般来说,当丘脑中继神经元处于突发模式时,脑电图是同步的,而当它们处于强直模式时,脑电图是不同步的。 [21]刺激 ARAS 通过抑制慢皮质波(0.3-1 Hz), δ波(1–4 Hz)和主轴波振荡(11–14 Hz)并通过提升伽玛波段(20–40 Hz)振荡。 [10]
从深度睡眠状态到清醒状态的生理变化是可逆的,并由 ARAS 介导。 [22]下丘脑的腹外侧视前核(VLPO) 抑制负责清醒状态的神经回路,VLPO 激活有助于睡眠开始。 [23]在睡眠期间,ARAS 中的神经元的放电率会低得多;相反,他们在清醒状态下的活动水平会更高。 [24]为了使大脑能够睡眠,必须通过抑制 ARAS 来减少到达皮质的上行传入活动。 [22]
ARAS 的临床意义
脑干ARAS 核团的肿块病变可导致意识水平的严重改变(例如昏迷)。 [25]中脑网状结构的双侧损伤可能导致昏迷或死亡。 [26]
直接电刺激 ARAS 会在猫中产生疼痛反应,并引发人类对疼痛的口头报告。</link>猫的上行网状结构激活会导致瞳孔散大, [27]这可能是由于长期疼痛引起的。这些结果表明 ARAS 回路与生理疼痛通路之间存在某种关系。 [27]
病理学
ARAS 的一些病理可能归因于年龄,因为随着年龄的增长,ARAS 的反应性似乎普遍下降。 [28]电耦合[upper-alpha 3]的变化被认为可以解释 ARAS 活动的一些变化:如果耦合下调,则高频同步(伽马带)也会相应减少。相反,上调的电耦合会增加快节奏的同步,从而导致觉醒和快速眼动睡眠驱动力的增加。 [30]具体来说,ARAS 的破坏与以下疾病有关:
发育影响
有几个潜在因素可能会对上行网状激活系统的发育产生不利影响:
网状脊髓下降束

网状脊髓束,也称为下降或前网状脊髓束,是锥体外系运动束,从网状结构下降[35]分为两条束,作用于供应躯干和近端肢体屈肌和伸肌的运动神经元。网状脊髓束主要参与运动和姿势控制,尽管它们也具有其他功能。 [36]下降网状脊髓束是通往脊髓的肌肉骨骼活动的四个主要皮质通路之一。网状脊髓束与其他三个通路一起协调运动控制,包括精细的操作。 [35]这四个通路可以分为两个主要系统通路——内侧系统和外侧系统。内侧系统包括网状脊髓通路和前庭脊髓通路,该系统提供姿势控制。皮质脊髓束和红核脊髓束通路属于侧向系统,提供对运动的精细控制。 [35]
内侧束和外侧束
该下降束分为两部分,内侧(或脑桥)和外侧(或髓质)网状脊髓束(MRST 和 LRST)。
- 内侧网状脊髓束负责兴奋反重力伸肌。该束的纤维源自尾部桥脑网状核和口腔桥脑网状核,并投射到脊髓的第七层和第八层。
- 外侧网状脊髓束负责抑制兴奋性轴向伸肌的运动。它还负责自动呼吸。该束的纤维来自髓质网状结构,大部分来自巨细胞核,并沿侧柱前部的脊髓长度下降。该束大部分终止于第七层,一些纤维终止于脊髓的第九层。
向相反方向传递信息的上升感觉束被称为脊髓网状束。
网状脊髓束的临床意义
网状脊髓束提供了下丘脑控制交感胸腰椎流出和副交感骶骨流出的通路。</link>
两个主要的下行系统将信号从脑干和小脑传送到脊髓,可以触发平衡和定向的自动姿势反应:来自前庭核的前庭脊髓束和来自脑桥和延髓的网状脊髓束。这些束的损伤会导致严重的共济失调和姿势不稳定。 [38]
脑干的物理或血管损伤,断开红核(中脑)和前庭核(脑桥)的连接,可能会导致去大脑强直,其神经系统症状是肌张力增加和牵张反射过度活跃。为了应对令人震惊或痛苦的刺激,手臂和腿都会伸展并向内转动。其原因是外侧前庭脊髓束和网状脊髓束的强直活动刺激伸肌运动神经元,而不受红核脊髓束的抑制。 [39]
红核水平以上的脑干损伤可能会导致皮质僵硬。当受到惊吓或疼痛的刺激时,手臂会弯曲,腿会伸展。原因是红核通过红核脊髓束抵消了来自外侧前庭脊髓束和网状脊髓束的伸肌运动神经元的兴奋。由于红核脊髓束仅延伸至颈脊髓,因此它主要通过兴奋屈肌和抑制伸肌来作用于手臂,而不是腿部。 [39]
历史
“网状结构”一词是由Otto Deiters在 19 世纪末创造的,与Ramon y Cajal的神经元学说不谋而合。艾伦·霍布森(Allan Hobson)在他的《重新审视网状结构》一书中指出,这个名字是神经科学中聚合场论衰落时代的词源遗迹。 “网状结构”一词的意思是“网状结构”,乍一看就是网状结构的样子。它被描述为要么太复杂而无法研究,要么是大脑的未分化部分,根本没有组织。埃里克·坎德尔(Eric Kandel)将网状结构描述为以与脊髓中间灰质类似的方式组织。这种混乱、松散且复杂的组织形式使得许多研究人员无法进一步研究大脑的这一特定区域。</link>这些细胞缺乏清晰的神经节边界,但具有清晰的功能组织和不同的细胞类型。除了泛泛而谈之外,“网状结构”一词已很少使用。现代科学家通常指的是构成网状结构的单个核。</link>
Moruzzi和Magoun于 1949 年首次研究了调节大脑睡眠-觉醒机制的神经成分。生理学家提出,大脑深处的某些结构控制着精神的觉醒和警觉性。 [20]人们曾认为,觉醒仅取决于大脑皮层对传入(感觉)刺激的直接接收。
由于直接对大脑进行电刺激可以模拟皮层电中继,马古恩利用这一原理在猫脑干的两个不同区域上演示了如何从睡眠中醒来。他首先刺激了上升的躯体和听觉路径;第二,一系列“从下脑干的网状结构通过中脑被盖、底丘脑和下丘脑到达内囊的上行中继”。 [40]后者特别令人感兴趣,因为这一系列中继不对应于任何已知的觉醒信号转导解剖通路,并被创造为上升网状激活系统(ARAS)。
接下来,通过在中脑前部的内侧和外侧部分放置损伤来评估这个新发现的中继系统的重要性。 ARAS 中脑中断的猫进入深度睡眠并显示出相应的脑电波。以另一种方式,上行听觉和躯体通路受到类似干扰的猫表现出正常的睡眠和觉醒,并且可以通过物理刺激唤醒。因为这些外部刺激在到达皮层的途中会被中断所阻碍,这表明上行传输必须通过新发现的 ARAS。
最后,马贡记录了脑干内侧部分的电位,发现听觉刺激直接激发了网状激活系统的部分区域。此外,坐骨神经的单次电击刺激也激活了内侧网状结构、下丘脑和丘脑。 ARAS 的兴奋不依赖于通过小脑回路的进一步信号传播,因为在去小脑和去皮后获得了相同的结果。研究人员提出,中脑网状结构周围的一列细胞接收来自脑干所有上升束的输入,并将这些传入信号传递到皮质,从而调节觉醒。 [40] [22]
参见
- 蓝斑
- 桥脚核
- 脑桥内侧网状结构
- 中脑网状结构
参考
- Gray, Henry. . Bartleby.com. [2019-09-12]. (原始内容存档于2018-04-21).
- . [2024-01-31]. (原始内容存档于2024-01-31).
- Iwańczuk W, Guźniczak P. . Anaesthesiol Intensive Ther. 2015, 47 (2): 162–167. PMID 25940332. doi:10.5603/AIT.2015.0015.
The ascending reticular activating system (ARAS) is responsible for a sustained wakefulness state. It receives information from sensory receptors of various modalities, transmitted through spinoreticular pathways and cranial nerves (trigeminal nerve – polymodal pathways, olfactory nerve, optic nerve and vestibulocochlear nerve – monomodal pathways). These pathways reach the thalamus directly or indirectly via the medial column of reticular formation nuclei (magnocellular nuclei and reticular nuclei of pontine tegmentum). The reticular activating system begins in the dorsal part of the posterior midbrain and anterior pons, continues into the diencephalon, and then divides into two parts reaching the thalamus and hypothalamus, which then project into the cerebral cortex (Fig. 1). The thalamic projection is dominated by cholinergic neurons originating from the pedunculopontine tegmental nucleus of pons and midbrain (PPT) and laterodorsal tegmental nucleus of pons and midbrain (LDT) nuclei [17, 18]. The hypothalamic projection involves noradrenergic neurons of the locus coeruleus (LC) and serotoninergic neurons of the dorsal and median raphe nuclei (DR), which pass through the lateral hypothalamus and reach axons of the histaminergic tubero-mamillary nucleus (TMN), together forming a pathway extending into the forebrain, cortex and hippocampus. Cortical arousal also takes advantage of dopaminergic neurons of the substantia nigra (SN), ventral tegmenti area (VTA) and the periaqueductal grey area (PAG). Fewer cholinergic neurons of the pons and midbrain send projections to the forebrain along the ventral pathway, bypassing the thalamus [19, 20].
- Brudzynski SM. . Journal of Molecular Neuroscience. July 2014, 53 (3): 436–445. PMID 24272957. S2CID 14615039. doi:10.1007/s12031-013-0179-1.
Understanding of arousing and wakefulness-maintaining functions of the ARAS has been further complicated by neurochemical discoveries of numerous groups of neurons with the ascending pathways originating within the brainstem reticular core, including pontomesencephalic nuclei, which synthesize different transmitters and release them in vast areas of the brain and in the entire neocortex (for review, see Jones 2003; Lin et al. 2011). They included glutamatergic, cholinergic, noradrenergic, dopaminergic, serotonergic, histaminergic, and orexinergic systems (for review, see Lin et al. 2011). ... The ARAS represented diffuse, nonspecific pathways that, working through the midline and intralaminar thalamic nuclei, could change activity of the entire neocortex, and thus, this system was suggested initially as a general arousal system to natural stimuli and the critical system underlying wakefulness (Moruzzi and Magoun 1949; Lindsley et al. 1949; Starzl et al. 1951, see stippled area in Fig. 1). ... It was found in a recent study in the rat that the state of wakefulness is mostly maintained by the ascending glutamatergic projection from the parabrachial nucleus and precoeruleus regions to the basal forebrain and then relayed to the cerebral cortex (Fuller et al. 2011). ... Anatomical studies have shown two main pathways involved in arousal and originating from the areas with cholinergic cell groups, one through the thalamus and the other, traveling ventrally through the hypothalamus and preoptic area, and reciprocally connected with the limbic system (Nauta and Kuypers 1958; Siegel 2004). ... As counted in the cholinergic connections to the thalamic reticular nucleus ...
- Schwartz MD, Kilduff TS. . The Psychiatric Clinics of North America. December 2015, 38 (4): 615–644. PMC 4660253
 . PMID 26600100. doi:10.1016/j.psc.2015.07.002.
. PMID 26600100. doi:10.1016/j.psc.2015.07.002. This ascending reticular activating system (ARAS) is comprised of cholinergic laterodorsal and pedunculopontine tegmentum (LDT/PPT), noradrenergic locus coeruleus (LC), serotonergic (5-HT) Raphe nuclei and dopaminergic ventral tegmental area (VTA), substantia nigra (SN) and periaqueductal gray projections that stimulate the cortex directly and indirectly via the thalamus, hypothalamus and BF.6, 12-18 These aminergic and catecholaminergic populations have numerous interconnections and parallel projections which likely impart functional redundancy and resilience to the system.6, 13, 19 ... More recently, the medullary parafacial zone (PZ) adjacent to the facial nerve was identified as a sleep-promoting center on the basis of anatomical, electrophysiological and chemo- and optogenetic studies.23, 24 GABAergic PZ neurons inhibit glutamatergic parabrachial (PB) neurons that project to the BF,25 thereby promoting NREM sleep at the expense of wakefulness and REM sleep. ... The Hcrt neurons project widely throughout the brain and spinal cord92, 96, 99, 100 including major projections to wake-promoting cell groups such as the HA cells of the TM,101 the 5-HT cells of the dorsal Raphe nuclei (DRN),101 the noradrenergic cells of the LC,102 and cholinergic cells in the LDT, PPT, and BF.101, 103 ... Hcrt directly excites cellular systems involved in waking and arousal including the LC,102, 106, 107 DRN,108, 109 TM,110-112 LDT,113, 114 cholinergic BF,115 and both dopamine (DA) and non-DA neurons in the VTA.116, 117
- Malenka RC, Nestler EJ, Hyman SE. 2nd. New York: McGraw-Hill Medical. 2009: 295. ISBN 978-0071481274.
The RAS is a complex structure consisting of several different circuits including the four monoaminergic pathways ... The norepinephrine pathway originates from the locus ceruleus (LC) and related brainstem nuclei; the serotonergic neurons originate from the raphe nuclei within the brainstem as well; the dopaminergic neurons originate in ventral tegmental area (VTA); and the histaminergic pathway originates from neurons in the tuberomammillary nucleus (TMN) of the posterior hypothalamus. As discussed in Chapter 6, these neurons project widely throughout the brain from restricted collections of cell bodies. Norepinephrine, serotonin, dopamine, and histamine have complex modulatory functions and, in general, promote wakefulness. The PT in the brain stem is also an important component of the ARAS. Activity of PT cholinergic neurons (REM-on cells) promotes REM sleep. During waking, REM-on cells are inhibited by a subset of ARAS norepinephrine and serotonin neurons called REM-off cells.
- Squire L. 4th. Amsterdam: Elsevier/Academic Press. 2013: 1095. ISBN 978-0123858702.
- Saper CB, Fuller PM. . Current Opinion in Neurobiology. June 2017, 44: 186–192. PMC 5531075 . PMID 28577468. doi:10.1016/j.conb.2017.03.021.
Parabrachial and pedunculopontine glutamatergic arousal system
Retrograde tracers from the BF have consistently identified one brainstem site of input that is not part of the classical monoaminergic ascending arousal system: glutamatergic neurons in the parabrachial and pedunculopontine nucleus ... Juxtacellular recordings from pedunculopontine neurons have found that nearly all cholinergic neurons in this region, as well as many glutamatergic and GABAergic neurons, are most active during wake and REM sleep [25], although some of the latter neurons were maximally active during either wake or REM, but not both. ... [Parabrachial and pedunculopontine glutamatergic neurons] provide heavy innervation to the lateral hypothalamus, central nucleus of the amygdala, and BF - Pedersen NP, Ferrari L, Venner A, Wang JL, Abbott SG, Vujovic N, Arrigoni E, Saper CB, Fuller PM. . Nature Communications. November 2017, 8 (1): 1405. Bibcode:2017NatCo...8.1405P. PMC 5680228 . PMID 29123082. doi:10.1038/s41467-017-01004-6.
Basic and clinical observations suggest that the caudal hypothalamus comprises a key node of the ascending arousal system, but the cell types underlying this are not fully understood. Here we report that glutamate-releasing neurons of the supramammillary region (SuMvglut2) produce sustained behavioral and EEG arousal when chemogenetically activated.
- Burlet S, Tyler CJ, Leonard CS. . J. Neurosci. April 2002, 22 (7): 2862–2872. PMC 6758338 . PMID 11923451. doi:10.1523/JNEUROSCI.22-07-02862.2002.
- Malenka RC, Nestler EJ, Hyman SE. 2nd. New York: McGraw-Hill Medical. 2009: 295. ISBN 978-0071481274.
Orexin neurons are located in the lateral hypothalamus. They are organized in a widely projecting manner, much like the monoamines (Chapter 6), and innervate all of the components of the ARAS. They excite the REM-off monoaminergic neurons during wakefulness and the PT cholinergic neurons during REM sleep. They are inhibited by the VLPO neurons during NREM sleep.
- Cherasse Y, Urade Y. . International Journal of Molecular Sciences. November 2017, 18 (11): 2334. PMC 5713303 . PMID 29113075. doi:10.3390/ijms18112334 .
The regulation of sleep and wakefulness involves many regions and cellular subtypes in the brain. Indeed, the ascending arousal system promotes wakefulness through a network composed of the monaminergic neurons in the locus coeruleus (LC), histaminergic neurons in the tuberomammilary nucleus (TMN), glutamatergic neurons in the parabrachial nucleus (PB) ...
- Fuller PM, Fuller P, Sherman D, Pedersen NP, Saper CB, Lu J. . The Journal of Comparative Neurology. April 2011, 519 (5): 933–956. PMC 3119596 . PMID 21280045. doi:10.1002/cne.22559.
- Kinomura S, Larsson J, Gulyás B, Roland PE. . Science. January 1996, 271 (5248): 512–515. Bibcode:1996Sci...271..512K. PMID 8560267. S2CID 43015539. doi:10.1126/science.271.5248.512.
This corresponds to the centro-median and centralis lateralis nuclei of the intralaminar group
- VandenBos, Gary R (编). . APA dictionary of psychology 2nd (Washington, DC: American Psychological Association). 2015: 57. ISBN 978-1433819445. doi:10.1037/14646-000.
a state of motor nonresponsiveness in nonhuman animals, produced by stroking, salient stimuli, or physical restraint. It is called “hypnosis” because of a claimed resemblance to human hypnosis and trance
- Svorad D. . Science. January 1957, 125 (3239): 156. Bibcode:1957Sci...125..156S. PMID 13390978. doi:10.1126/science.125.3239.156.
- Jang SH, Kwon HG. . Neurosci. Lett. October 2015, 606: 200–203. PMID 26363340. S2CID 37083435. doi:10.1016/j.neulet.2015.09.004.
- Jones, BE. . Annals of the New York Academy of Sciences. 2008, 1129 (1): 26–34. Bibcode:2008NYASA1129...26J. PMID 18591466. S2CID 16682827. doi:10.1196/annals.1417.026.
- Purves et al (2018b),Box 28A – Electroencephalography, pp. 647–649
- Steriade, M. . Science. 1996, 272 (5259): 225–226. Bibcode:1996Sci...272..225S. PMID 8602506. S2CID 39331177. doi:10.1126/science.272.5259.225.
- Reiner, P. B. . Seminars in Neuroscience. 1995, 7 (5): 355–359. S2CID 5575547. doi:10.1006/smns.1995.0038.
- Evans, B.M. . Neurophysiologie Clinique. 2003, 33 (1): 1–10. PMID 12711127. S2CID 26159370. doi:10.1016/s0987-7053(03)00002-9.
- Purves et al (2018b)
- Mohan Kumar V, Mallick BN, Chhina GS, Singh B. . Exp. Neurol. October 1984, 86 (1): 40–52. PMID 6479280. S2CID 28688574. doi:10.1016/0014-4886(84)90065-7.
- Tindall SC. https://www.ncbi.nlm.nih.gov/books/NBK380/#A1740
|chapterurl=缺少标题 (帮助). . Butterworth Publishers. 1990 [2008-07-04]. ISBN 978-0409900774. (原始内容存档于2009-01-29). - Nolte (编). 5th. : 262–290.
- Ruth RE, Rosenfeld JP. . Exp. Neurol. October 1977, 57 (1): 41–56. PMID 196879. S2CID 45019057. doi:10.1016/0014-4886(77)90043-7.
- Robinson, D. . Clinical Neurophysiology. 1999, 110 (8): 1427–1434. PMID 10454278. S2CID 38882496. doi:10.1016/S1388-2457(99)00078-4.
- Lawrence, Eleanor (编). . Henderson's dictionary of biology 13th (Pearson Education Limited). 2005: 195. ISBN 978-0131273849.
- Garcia-Rill E, Heister DS, Ye M, Charlesworth A, Hayar A. . Sleep. 2007, 30 (11): 1405–1414. PMC 2082101 . PMID 18041475. doi:10.1093/sleep/30.11.1405.
- Schwartz JR, Roth T. . Curr Neuropharmacol. December 2008, 6 (4): 367–378. PMC 2701283 . PMID 19587857. doi:10.2174/157015908787386050.
- Vincent, S. R. . Journal of Chemical Neuroanatomy. 2000, 18 (1–2): 23–30. PMID 10708916. S2CID 36236217. doi:10.1016/S0891-0618(99)00048-4.
- Hall RW, Huitt TW, Thapa R, Williams DK, Anand KJ, Garcia-Rill E. . Clin Neurophysiol. June 2008, 119 (6): 1281–1291. PMC 2670248 . PMID 18372212. doi:10.1016/j.clinph.2007.12.021.
- Garcia-Rill E, Buchanan R, McKeon K, Skinner RD, Wallace T. . Neurotoxicology. September 2007, 28 (5): 915–923. PMC 3320145 . PMID 17368773. doi:10.1016/j.neuro.2007.01.007.
- Squire L. 4th. Amsterdam: Elsevier/Academic Press. 2013: 631–632. ISBN 978-0123858702.
- FitzGerald MT, Gruener G, Mtui E. . Philadelphia: Saunders Elsevier. 2012: 192. ISBN 978-0702037382.
- Brownstone, Robert M.; Chopek, Jeremy W. . Frontiers in Neural Circuits. 2018, 12: 30. ISSN 1662-5110. PMC 5915564 . PMID 29720934. doi:10.3389/fncir.2018.00030 .
- Pearson, Keir G; Gordon, James E. Kandel , 编. 5th. United States: McGraw-Hill. 2013. The Brain Stem and Cerebellum Integrate Sensory Signals for Posture, p. 954. ISBN 978-0071390118. Editors list列表中的
|first2=缺少|last2=(帮助); Editors list列表中的|first4=缺少|last4=(帮助) - Michael-Titus et al (2010b)
- Magoun HW. . AMA Arch Neurol Psychiatry. February 1952, 67 (2): 145–154; discussion 167–171. PMID 14893989. doi:10.1001/archneurpsyc.1952.02320140013002.
其他参考资料
外部链接
 维基词典中的词条「reticular formation」
维基词典中的词条「reticular formation」
- Animal hypnosis is a state in nonhuman animals in which there is no motor response. The state may occur because of stroking, salient stimuli, or physical restraint. The name comes from its claimed resemblance to human hypnosis and trance.[15]
- An EEG's electrode on the scalp measures the activity of a very large number of pyramidal neurons in the underlying brain region. Each neuron generates a small electrical field that changes over time. In the sleep state, the neurons activate at approximately the same time, and the EEG wave, representing the summation of the neurons' electrical fields, tend to be in phase and has higher amplitude, and hence it is "synchronized." In the waking state, they do not activate at the same time because of irregular or out-of-phase inputs, the EEG wave, representing the algebraic sum, will have a smaller amplitude, and hence "dysynchronized."[19]
- Electrical coupling is the passive flow of electric current from one cell into an adjacent cell through gap junctions, such as the cells in the heart muscle or the neurons with electrical synapses. Electrically coupled cells fire synchronously because generated currents in one cell rapidly spread to the other cells.[29]