青島龍屬
青島龍(屬名:,發音:/tʃɪŋdaʊˈsɔːrəs/,意為「青島的蜥蜴」;tsintao為舊式拼音)[1]是一屬具有頭冠的鴨嘴龍科恐龍,生存於白堊紀晚期的東亞,相當於現在的中國山東省,化石發現於萊陽市附近、屬於坎潘階的王氏群金剛口組地層。模式種棘鼻青島龍(T. spinorhinus)由中國古生物學家楊鍾健於1958年敘述、命名。正模標本頭骨保存了一根長4公分的刺狀突起,過去被認為這構成一根如獨角獸般的獨特角狀頭冠,但近期研究發現這個刺只是頭冠斷裂剩下的一小部分。
| 青岛龙属 化石时期:晚白堊世 | |
|---|---|
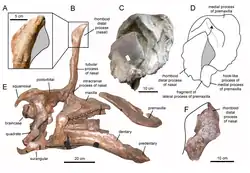 | |
| 正模標本的頭骨與斷掉的頭冠 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | †鸟臀目 Ornithischia |
| 亚目: | †鸟脚亚目 Ornithopoda |
| 科: | †鴨嘴龍科 Hadrosauridae |
| 亚科: | †賴氏龍亞科 Lambeosaurinae |
| 族: | †青岛龙族 Tsintaosaurini |
| 属: | †青岛龙属 Tsintaosaurus Young, 1958 |
| 模式種 | |
| †棘鼻青島龍 Tsintaosaurus spinorhinus Young, 1958 | |
發現及命名

1950年,在山東省萊陽市金剛口鎮附近的溪口地區發現了數具大型鴨嘴龍類遺骸。1958年,這些化石由中國古生物學家楊鍾健命名、敘述為模式種棘鼻青島龍(T. spinorhinus),屬名取自發現地附近的青島市,但採用的是當時普遍的威妥瑪拼音;種名將拉丁語的spina(棘刺)與希臘語的/rhis(鼻子)組合而成,意為「棘刺般的鼻子」,象徵著當時認為具有獨特的刺狀頭冠。[2]
青島龍的化石皆出土於王氏群的金剛口組地層,年代可追溯至晚白堊世坎潘階,約7200萬年前。正模標本IVPP AS V725是一個帶有頭骨的部分骨骼;副模標本IVPP V818則是個頭蓋骨。在同一地區,還挖掘出大量關節脫落的部分骨骼,其中一些被楊歸入青島龍,其他則由楊於1958年命名為金剛口譚氏龍(Tanius chingkankouensis)以及曾於1976年命名為萊陽譚氏龍(Tanius laiyangensis)。到了現代後兩個物種通常被認為是次異名或疑名。未來的研究可能會有大量的額外標本歸入青島龍。
敘述
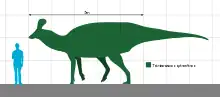

青島龍身長約8.3公尺、體重約2.5公噸。[3]作為一種鴨嘴龍科,青島龍具有典型的似鴨吻部,以及大量強壯牙齒所構成的齒系,適合咀嚼植物。平常主要以四足行走,但也可用二足方式逃離掠食動物。如同其他鴨嘴龍類,青島龍可能以群體方式共同生活。
頭冠
青岛龙的头冠起初被重建成独角兽的形状。其保存下来的是根长约40公分的突起物,从吻部後部顶端几乎垂直地突出。头冠构造中空、上端似乎呈現分叉狀,而沒有任何相关物种有類似的頭冠,牠們通常都是長著葉瓣状的头冠。1990年,韋尚保和霍納对青岛龙头冠的存在表示怀疑,他們認為这其实是从口鼻顶部碎裂,并在化石过程中被向上扭曲的鼻骨;他们更提出青岛龙其实缺乏這個具鑑別性的头冠特徵,因此可能是谭氏龙的异名。然而巴菲陶等人於1993年在对青島龍頭骨進行重新檢驗后,得出的結論為:头冠並非變形或位移、也不是人工復原的產物。除此之外,已发现的第二具标本也有着类似的垂直構造,表明了青岛龙的头冠確實存在且為独立的属。[4]

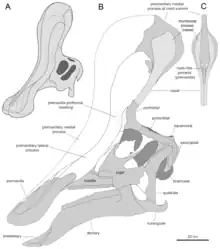


2013年,皮耶妥-馬奎茲(Albert Prieto-Márquez)和華格納(Jonathan Wagner)根據對IVPP V829標本(一個屬於青島龍的前上頜骨)的鑑定提出了新版重建圖,表示獨角獸形狀的突起只是大型頭冠的後方殘留部分,而頭冠前部是從吻部前端的前上頜骨所延伸出來的。这些擴張菱形接触面以及鼻骨頂部突起的擴張構成了头冠的后部。头冠的后基部被前额骨的附生物所覆盖。癒合的鼻骨形成了中空的管状结构。沿著方骨的位置來看,头冠高度將超越头骨后部的高度。頭冠大致上是垂直向,頂部稍微向後;而正模标本的头冠向前倾斜则是化石变形的结果。[5]
皮耶妥-馬奎茲和華格納的新版重建也延伸出關於頭冠內部空氣管道的新推論。楊曾推測正模標本保存的空心管是主要的空氣吸入口,但皮耶妥-馬奎茲和華格納反駁道這個管子下方是封閉的構造,而通常賴氏龍亞科的氣管位置會在更前面,其骨質鼻孔被前上頜骨完全蓋住。他們推測青島龍具有典型的賴氏龍亞科吻部配置,當吸氣時,空氣透過上嘴喙後方前上頜骨的成對「假鼻孔」(pseudonares)進入頭骨,再經過前上頜骨中突下方的成對氣管來到頭冠頂部,最後在管狀構造的共通腔室匯合。腔室後部由鼻骨構成,可能對應著實際鼻腔的位置。鼻腔被前上頜骨中突在周圍構成的鈎狀構造分隔成兩個較小的腔室,一前一後。空氣從後面的腔室傳送到下方往顱腔內部移動。雖然普遍認為只有一條管道有此功能,但皮耶妥-馬奎茲和華格納發現在鼻骨中,有一對朝下的管道通往前上頜骨側突內部的痕跡。據此他們得出結論為:整段氣流可能被分隔開來,中間的共通腔室可能也被軟骨隔膜區分為左右兩部分。[5]

後部鼻骨構成的管狀構造並非空氣管道的結論使皮耶妥-馬奎茲和華格納提出替代方案來解釋其功能。他們推測這種管狀結構有著相當的強韌性與較低的骨頭密度,可能可以用來減輕頭冠重量。青島龍的這項特徵與較進階的賴氏龍亞科有所差異,後者透過額骨向前延伸構成骨片來支撐頭冠。[5]
一份2017年的CT掃描研究指出,頭冠中的空腔結構純屬人為想像,且頭冠與頭骨表面的斷裂面無法吻合,因此難以證實各部位頭骨彼此的關聯性,也無法得知頭冠的確切形狀。[6]另一個2020年的研究表示確定上述的其中一個空腔不存在,因此頭冠的朝向應該更偏往前方,鼻骨和額骨之間交界處會有個前頭孔。[7]
分類
在賴氏龍亞科中,青島龍可能與歐洲的似凹齒龍和匙龍(後兩者可能是異名)構成一個演化支,稱為青島龍族。[8]
以下演化樹取自皮耶妥-馬奎茲和華格納(2013)的系統發生學分析,顯示出青島龍位於賴氏龍亞科中相對基礎的演化位置:[9]
| 赖氏龙亚科 Lambeosaurinae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
古生態學
一份關於王氏群連續地層的恐龍蛋研究顯示,青島龍所生活的金剛口組屬於恐龍多樣性較高的區塊,當地氣候會比下方較古老的將軍頂組來得乾燥一些。[10]
同地區出產的恐龍還包括:櫛龍亞科的山東龍、萊陽龍;角龍科的中國角龍;原始角龍類諸城角龍和坐骨角龍;蜥腳類的諸城巨龍;暴龍科的諸城暴龍。
青島龍的化石還沒在中國以外的地區發現過,但在蒙古的同時代地層出產過許多也同時出現在中國的物種或其近親,因此可推測青島龍的分布範圍可能比所知的更廣。
延伸閱讀
- 鴨嘴龍類研究歷史
參考來源
- Creisler, B. . DOL Dinosaur Omnipedia. 2002 [24 Feb 2010]. (原始内容存档于31 December 2005).
- Young, C.-C. . Palaeontologia Sinica, New Series C (Whole Number). 1958, 42 (16): 1–138.
- Gregory S. Paul. . United States: Princeton University Press. 2010: 308. ISBN 978-0-691-13720-9.
- Buffetaut, E.; Tong, H. . 2 317. C.R. Academy of Science Paris: 1255–1261. 1993.
- Prieto-Márquez, A.; Wagner, J. R. . PLOS ONE. 2013, 8 (11): e82268. Bibcode:2013PLoSO...882268P. PMC 3838384
 . PMID 24278478. doi:10.1371/journal.pone.0082268.
. PMID 24278478. doi:10.1371/journal.pone.0082268. - ZHANG Jia-Liang, WANG Qiang, JIANG Shun-Xing, CHENG Xin, LI Ning, QIU Rui, ZHANG Xin-Jun & WANG Xiao-Lin, 2017, "Review of historical and current research on the Late Cretaceous dinosaurs and dinosaur eggs from Laiyang, Shandong", Vertebrata PalAsiatica 55(2): 187-200
- Jialiang Zhang; Xiaolin Wang; Shunxing Jiang & Guobiao Li. 2020. "Internal morphology of nasal spine of Tsintaosaurus spinorhinus (Ornithischia: Lambeosaurinae) from the upper cretaceous of Shandong, China". Historical Biology: An International Journal of Paleobiology
- Prieto-Márquez, A.; and Wagner, J.R. . Cretaceous Research. 2009,. online preprint. doi:10.1016/j.cretres.2009.06.005.
- Prieto-Marquez, A.; Vecchia, F. M. D.; Gaete, R.; Galobart, A. . PLOS ONE. 2013, 8 (7): e69835. PMC 3724916 . PMID 23922815. doi:10.1371/journal.pone.0069835.
- Zhao, ZiKui; Zhang, ShuKang; Wang, Qiang; Wang, XiaoLin. . Chinese Science Bulletin. 2013, 58 (36): 4663–4669. Bibcode:2013ChSBu..58.4663Z. S2CID 131373599. doi:10.1007/s11434-013-6059-9.