ABC模型
被子植物花发育的ABC模型由恩里科·科恩和埃利奥特·迈耶罗维茨在1991年提出。[1]这个模型是以对花器官发育有缺陷的突变体的观察为基础的。ABC模型概括了在花的不同部位中,不同类型的转录因子是怎样起作用或不起作用,从而控制了花部器官的发育的。
模型的提出
两个关键发现导致了ABC模型的提出。第一,同源异型突变使在正常情况下应发育某种器官的部位发育出了另一种器官。比如野生蔷薇只有5枚花瓣和众多的雄蕊,然而,园艺蔷薇却具有一个同源异型基因,使一些本应发育成雄蕊的组织发育成了花瓣。第二,每一个能影响花部器官的决定的基因都可以同时影响两种花部器官,或者是影响花瓣和萼片,或者是影响花瓣和雄蕊。[2]
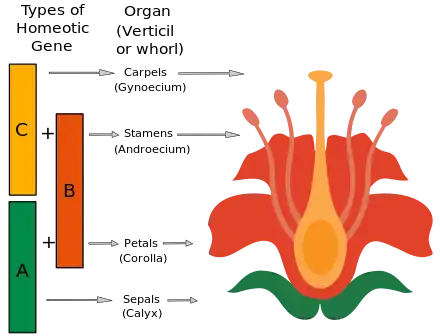
花部器官决定基因因而可以根据它们所影响的器官而分成三类。A类基因的突变影响萼片和花瓣。B类基因的突变影响花瓣和雄蕊。C类基因的突变则影响雄蕊和心皮。所有这三类基因都是可转录成蛋白质的同源异型基因。除其中一个基因(APETALA2)外,这些基因编码的蛋白质都含有一个MADS盒结构域,这一结构域可以结合含有特定基序(即CArG-box motif, CC[A/T]6GG)的DNA序列,从而在DNA转录时起到调控子的作用。据信,这些基因都是调控其他控制器官发育的基因的主控基因。
| 金鱼草 | 拟南芥 | |
|---|---|---|
| A类 | SQUAMOSA (SQUA) | APETALA2 (AP2) APETALA1 (AP1) |
| B类 | DEFICIENS (DEF) GLOBOSA (GLO) | APETALA3 (AP3) PISTILLATA (PI) |
| C类 | PLENA (PLE) FARINELLI (FAR) | AGAMOUS (AG) |
ABC模型认为,A基因本身决定萼片,A和B基因共同决定花瓣,B与C基因共同决雄蕊,C基因决定心皮。此外,A基因与C基因相互拮抗。当A基因(AP1、AP2)突变后,导致第一轮花器官中的花萼突变为心皮,第二轮花器官中的花瓣突变为雄蕊。B基因(AP3、PI)突变后,花萼替代了第二轮花器官中的花瓣,第三轮花器官中的雄蕊变为心皮。若C基因(AG)失活,则第三轮花器官中的雄蕊转变为花瓣,第四轮花器官中的心皮转变为花瓣。在ag突变体的心皮位置长出的花瓣实际上是一朵新的花。在AG的强突变体里,有时在花心位置可以长出好几层花瓣,这就是常见的玫瑰花重瓣表型的分子遗传基础。
小结:
E类功能的发现及ABC模型的扩展
2000年,Yanofsky实验室利用反向遗传学的手段研究了拟南芥中三个亲缘关系很近的横向同源基因AGL2,AGL4, AGL9三重突变体的功能,发现其中所有花器官都表现出萼片的特征,因此证明B和C类基因的功能同时需要这三个MADS-box基因的功能[3]。这三个基因被分别重新命名为为SEPALLATA1,SEP2和SEP3。2001年,日本京都大学研究人员Takashi Honma和Koji Goto发表文章证明SEP3可与A类,B类及C类基因的蛋白产物相互作用以形成三聚体或四聚体复合物,且相应基因的异位表达足以促使叶子发育成花器官的形态,为ABC模型及E类基因在花原基的形成和花器官特征的建立过程中的作用提供了分子层次的解释。[4]。Theißen等据此提出了“四聚体”模型[5] [6] 2004年,Yanofsky实验室确定MADS-box基因AGL3即第4个SEP基因,命名为SEP4,其功能与之前发现的3个SEP基因高度冗余[7]。 在这些研究的基础上,ABC模型发展为ABCE模型。
ABCE基因的调控及其下游作用因子
B基因在第四轮花器官中的表达受到SUPERMAN基因的抑制。sup突变体的表型为第四轮心皮数量大大减少。而雄蕊数量大大增加。如前三轮花器官中的基因发育正常,只有第四轮花器官中有B基因异位表达,导致该轮花器官发育为雄蕊。大量单突变、双突变、三突变变体以及基因过量表达的研究结果均在不同程度上支持这个模型。“ABC”模型的提出时近年植物发育生物学研究中的一个非常重要的突破,可以解释多个基因在器官特征性发育中的作用。
ABC功能的保守性及模型的推广
ABC模型最初主要基于两个核心真双子叶植物拟南芥和金鱼草中的经典遗传结果总结提出。这一模型在其它基部双子叶植物以及单子叶植物中的适用性或保守性,不仅对于推广该模型的研究结果具有直接的指引作用,而且对于理解被子植物花器官的进化及物种多样性的形成具有基础性指导意义。在这方面,早期研究显示,B和C类基因及其功能在水稻,玉米等单子叶植物中具有显著的保守性。[8][6] B类基因的功能甚至在裸子植物中都有一定的保守性。在ABC模型中,A类基因及其功能被认为是最不保守的。其中,拟南芥A类基因AP2的功能保守性十分有限。拟南芥中的另一个A类基因,AP1,有一个序列高度近似的近缘同源基因CAULIFLOWER及一个亲缘关系较远的同源基因FUL,构成MADS-box基因家族的FUL/AP1分支。分子进化分析表明FUL/AP1分支在被子植物祖先或更早已经形成,在单子叶植物中可以找到明显的FUL/AP1分支的基因。但长期以来,研究者多认为单子叶植物中的FUL及其同源基因并不具有对应的A类功能。但是,近期的一项研究发现,水稻中FUL/AP1分支基因的功能,具有拟南芥中A类基因的典型特征,因此明确提出了单子叶植物中A类功能的保守性。[9]
基于对基部被子植物花器官形态及ABC基因表达模式的研究分析,研究者对ABC模型提出了不同版本的修正,以解释基部被子植物花器官形态的分子基础。其中,Soltis等于2007年提出的“模糊边界”模型,较为系统的总结了适用于基部被子植物的修正的ABC模型。[10]
参考文献
引用
- Coen, Henrico S.; Elliot M. Meyerowitz. . Nature. 1991, 353: 31–37. doi:10.1038/353031a0.
- Fosket, Donald E. . San Diego: Academic Press. 1994: 498-509. ISBN 0-12-262430-0.
- Pelaz, Soraya; Gary S. Ditta, Elvira Baumann, Ellen Wisman, Martin F. Yanofsky. . Nature. 2000, 405: 200–203. doi:10.1038/35012103.
- Honma, Takashi; Koji Goto. . Nature. 2001, 409: 525–529. doi:10.1038/35054083.
- Theißen, Günter; Heinz Saedler. . Nature. 2001, 409: 469–471. doi:10.1038/35054172.
- Goto, Koji; Junko Kyozuka, John L Bowman. . Current Opinion in Genetics & Development. 2001, 11 (4): 449–456. doi:10.1016/S0959-437X(00)00216-1.
- Ditta, Gary; Anusak Pinyopich, Pedro Robles, Soraya Pelaz, Martin F.Yanofsky. . Current Biology. 2004, 14 (21): 1935–1940. doi:10.1016/j.cub.2004.10.028.
- Ambrose, Barbara A.; David R. Lerner, Pietro Ciceri, Christopher M. Padilla, Martin F. Yanofsky, Robert J. Schmidt. . Molecular Cell. 2000, 5 (3): 569–579. doi:10.1016/S1097-2765(00)80450-5.
- Wu, Feng; Xiaowei Shi, Xuelei Lin, Yuan Liu, Kang Chong, Günter Theißen, Zheng Meng. . The Plant Journal. 2017, 89 (2): 310–324. doi:10.1111/tpj.13386.
- Soltis, Douglas; André S. Chanderbali, Sangtae Kim, Matyas Buzgo, Pamela S. Soltis. . Annals of Botany. 2005, 100 (2): 155–163. doi:10.1093/aob/mcm117.
书籍
- 朱玉贤、李毅、郑晓峰:《现代分子生物学》(第三版). 高等教育出版社. 2007.11:409-410