球囊菌門
球囊菌門(學名:Glomeromycota,AM fungi),又稱聚合菌門、鏽球菌門,是真菌界的一個門,目前約已發現400種物種。本門真菌可與陸生植物形成叢枝菌根,這種共生構造可協助植物吸收土壤中的無機鹽,尤其是磷的吸收,並據信是早期植物能適應陸地環境的重要關鍵[1]。超過八成的維管束植物都能與球囊菌產生共生關係,且在苔蘚植物等沒有真實根部構造的植物中亦有樹狀菌根,可見叢枝菌根在陸地生態系的重要性[2]。
| 球囊菌門 | |
|---|---|
 | |
| Lotus corniculatus var. japonicus(植物)和Gigaspora margarita(球囊菌)形成的菌根 | |
| 科学分类 | |
| 界: | 真菌界 Fungi |
| 亚界: | 毛黴亞界 Mucoromyceta |
| 门: | 球囊菌門 Glomeromycota |
| 亞門及綱 | |
| |
主要結構
.png.webp)
叢枝菌根原先稱為囊叢枝內生菌根(vesicular-arbuscular mycorrhiza, VAM) ,因菌絲能在感染的根部皮層細胞內形分支的叢枝狀體 (arbuscule) 以及在菌絲末端膨大形成囊泡(vesicle) 而得名,但近年來發現並非所有的叢枝菌根菌皆可於根內形成上述兩種構造, 因此已改稱為叢枝菌 (arbuscular mycorrhiza, AM) 。 Gigaspora 和Scutellospora 兩屬的叢枝菌根菌皆是將囊泡 (vesicle) 產生於根外土壤中。
泡囊體(Vesicles)
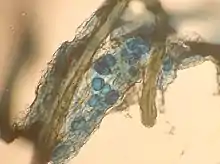
菌絲膨大而形成,儲存能量(lipid),不是每種內生菌根菌都有,形成於細胞內或細胞間。
叢枝狀體(Arbuscule)
為高度分支的特化菌絲構造,會穿透細胞壁進入植物細胞,但不破壞細胞膜,提供代謝與養分的雙向傳遞,所有內生菌根菌皆有,壽命短,形成後數天便分解。
厚膜孢子(Chlamydospore)
為菌根菌主要的無性繁殖構造,具有一到多層壁,多個細胞核,可殘存於土中數年,依其形成方式與壁層構造,做為內生菌根菌形態鑑定的主要依據。
共生關係

聚合菌可構成陸生植物的叢枝菌根,也因此被稱為「AM Fungi」(Arbuscular mycorrhiza)[2],可在植物的根中形成叢枝狀體,作為養分的交換構造。許多證據顯示在此共生關係中,聚合菌是絕對共生菌,須依賴植物合成的碳及能量才能生存,但有對某些物種的研究表明聚合菌可能可以單獨存在[3]。形成菌根的聚合菌廣泛的分布在全世界的土壤中,和植物的根(或苔蘚植物的假根)形成共生關係,它們也可以生存在鹽度很高的沼澤中,和附生植物形成共生關係。土管菌則是少數不與植物形成菌根的聚合菌,而是與一種稱為点形念珠藻的藍綠菌形成內共生關係,在此共生關係中,球囊菌形成多核的細胞,並將念珠藻屬的藍綠菌「吞入」。過去常以「Geosiphon pyrifome」稱呼此內共生體,但現在這個名稱只用來專指真菌,內共生體改以「Geosiphon symbiosis」稱呼。聚合菌-念珠藻的共生關係是目前發現的唯一一種真菌和藍綠菌的內共生關係,它不屬於地衣,因為它們涉及細胞內的連結,且就結構與演化上的證據而言這種關係比較類似叢枝菌根[2][4]。
在球囊菌-念珠藻共生關係中,還有一類功能不明、類似細菌的有機物(bacteria like organisms),由於這類有機物出現在球囊菌門的各分枝物種中,而被認為可能在演化的早期就已和聚合菌形成共生關係,並進一步演變成內共生[2][5]。
在整個系統中,念珠藻可獨立生存,所以球囊菌是義務性共生者,不過聚合菌除了與念珠藻形成叢枝菌根外,也可與植物形成樹狀菌根,這個說法尚未獲得證實,但已有若干證據支持[2]。
生殖
球囊菌門的菌絲體是多核的,通常採用無性生殖,在菌絲的末端產生直徑約80~500微米[6]的孢子(稱為Glomerospores)。在某些情況中,孢子生成後會進一步形成球囊狀的構造(saccule)[7]。
種系發生學
早期對於球囊菌門的研究多半著重在其型態及土壤中、菌根內外孢子堆的構造[8]「球囊菌門」的概念最早是在原囊黴屬(Acaulospora)及Gigaspora兩個屬被提出,並和接合菌門Endogonaceae下的Glomus屬[9][10]共同被提升為科,合為球囊黴目(屬於接合菌門)[11]。1998年Cavalier與Smith提出「球囊菌綱(Glomomycetes)」的概念,並將球囊黴目歸入其中(仍屬接合菌門)[12]。後來經Schüßler對該目下部分物種小亞基rRNA(SSUrRNA)的分析發現球囊菌目可自組一個門,便將其由接合菌門移出,新成立「球囊菌門」,又因發現有能產生類似球囊黴目孢子的種類應該屬於另一個更分歧的種系[13],而新增了類球囊霉目(Paraglomerales)、原囊黴目(Archaeosporales)與多孢囊霉目(Diversisporales)三個目[7],與念珠藻(Nostoc punctiforme)形成叢枝菌根的Geosiphon pyriformis即屬於原囊黴目。
現在的分類即是透過分子生物學的分析,不斷修正才定出的。對於本門物種的分類研究尚未完成,有待繼續研究,例如有些研究認為Glomus屬下的許多物種應被分到其他科[6]或屬[14]比較合適。
有研究分析各種真菌rDNA的序列,發現球囊菌門在演化上是雙核亞界的姊妹群[7]。
球囊菌門的化石種包括生存於志留紀晚期至泥盆紀晚期的原杉菌。
參考資料
- Brundrett, M.C. . New Phytologist. 2002, 154: 275–304. doi:10.1046/j.1469-8137.2002.00397.x.
- A. Schüßler. . Schuessler Lab. 2008. (原始内容存档于2011-07-19) (英语).
- Hempel, S., Renker, C. & Buscot, F. . Environmental Microbiology. 2007, 9 (8): 1930–1938. PMID 17635540. doi:10.1111/j.1462-2920.2007.01309.x.
- 吳繼光, 林素禎. . J. Taiwan Agric. Res.. 2016, 65 (3): 238–260. doi:10.6156/JTAR/2016.06503.02.
- Maria Naumann, Arthur Schüßler and Paola Bonfante. . The ISME Journal 4, 862-871 (July 2010). [2011-06-07]. doi:10.1038/ismej.2010.21. (原始内容存档于2016-08-26).
- Simon, L., Bousquet, J., Levesque, C., Lalonde, M. . Nature. 1993, 363 (6424): 67–69. doi:10.1038/363067a0.
- Schüßler, A.; et al. . Mycol. Res. December 2001, 105 (12): 1413–1421 [2011-06-10]. doi:10.1017/S0953756201005196. (原始内容存档于2011-05-24).
- Tulasne, L.R., & C. Tulasne. . Giornale Botanico Italiano. 1844, 2: 55–63.
- Thaxter, R. . Proc. Am. Acad. Arts Sci. 1922, 57 (12): 291–341 [2011-06-10]. doi:10.2307/20025921. (原始内容存档于2021-10-27).
- J.W. Gerdemann & J.M. Trappe. . Mycologia Memoirs. 1974, 5: 1–76.
- J.B. Morton & G.L. Benny. . Mycotaxon. 1990, 37: 471–491 [2011-06-10]. (原始内容存档于2021-08-09).
- Cavalier-Smith, T. . Biol. Rev. Camb. Philos. Soc. 1998, 73: 246. (as "Glomomycetes")
- Redeker, D. . Plant and Soil. 2002, 244: 67–73. doi:10.1023/A:1020283832275.
- Walker, C. . Agronomie. 1992, 12: 887–897. doi:10.1051/agro:19921026.
外部連結
| 維基物種上的相關:球囊菌門 |
- Tree of Life Glomeromycota (页面存档备份,存于)
- Glomeromycota at the International Culture Collection of VA Mycorrhizal Fungi (INVAM)
- Glomeromycota at the University of Sydney Fungal Biology s|ite
- 'AMF-phylogeny' (页面存档备份,存于) - 'Glomeromycota database' web-site at the University of Munich