光合作用
光合作用也称光能合成(英語:),是许多食物网基层的等生產者(自养生物)利用光能将水、二氧化碳或硫化氫等无机物转变成可以储存化学能的有机物(比如碳水化合物)的生物过程[1]。根据化学反应所产生的副产品,光合作用可分为產氧光合作用(oxygenic photosynthesis)和不產氧光合作用(anoxygenic photosynthesis)两类,分别使用不同的感光色素(photopigment),而且會因為不同環境改變反應速率。通俗意义上的“光合作用”主要指产氧光合作用。

自元古宙开始,地球生物圈主要以产氧光合作用为主。进行此类光合作用的生物(比如各种植物、绿藻和蓝绿菌)主要依赖卟啉衍生物——叶绿素作为反应中心的感光色素,主要吸收可见光谱中的红色和蓝色波段(不吸收绿光,因此呈现绿色),并以此将水和二氧化碳转换为碳水化合物和氧气,其理论最高能量轉換效率約為4.6-6%,实际效率仅有1.9-2.5%[2][3]。作为起始端的生产者,植物和藻类通过光合作用生产的有机物可以被其它异养生物摄入并消化,然后通过食物链将生物能传递给生态系统内的其它消费者。對大多數生物來説,這個過程是賴以生存的關鍵。而地球上的碳循环,光合作用是其中最重要的一环。
除此之外,许多厌氧微生物(比如绿硫细菌、紫细菌、酸杆菌和太阳杆菌)会进行不产氧光合作用,主要用菌绿素分解硫化氢获取氢正离子和二氧化碳发生反应,并产生碳水化合物和硫。除此之外,盐杆菌等古菌会用视黄醛和其视紫质衍生物来吸收绿光(因此呈现紫色)驱动跨膜蛋白中的离子泵并直接生产三磷酸腺苷和化学能,并不进行固碳反应。以视黄醛为基础的光合作用很可能在太古宙早期就已经出现,是地球史上最早的一种利用日光生产有机物的生化现象(参见紫色地球假说)。
研究历史


虽然一些在光合作用中的步骤仍不能被完全理解,但是整体的光合方程式自19世纪以来是已知的。
- 古希腊哲学家亚里士多德认为,植物生长所需的營養全来自土中。
- 1642年比利时人范·海尔蒙特做了「柳树实验」,推論植物的重量主要不是来自土壤而是来自水。但他没有發現空气中的物质也参与了有机物的形成。
- 1771年,英国的普里斯特利发现植物能够更新由于蜡烛燃烧或动物呼吸而变得污浊了的空气;但他并没有发现光的重要性。普里斯特利还发现置於密封玻璃罩內的老鼠極易窒息,但是如果加入一片新鮮薄荷葉,老鼠就可以甦醒。
- 1773年,荷兰的英格豪斯证明只有植物的绿色部分在光下才能起使空气变“好”的作用。
- 1774年,英國的普里斯特利發現綠色的植物會製造、釋放出氧氣。
- 1782年,瑞士的瑟訥比埃發現,即使植物有受到陽光照射,照樣會釋放出氧氣。
- 1804年,瑞士的索绪尔通过定量研究进一步证实二氧化碳和水是植物生长的原料。
- 1845年,德国的迈尔发现植物把太阳能转化成了化学能。[4]
- 1864年,德国的萨克斯发现光合作用会产生淀粉。
- 1880年,德国的恩格尔曼发现叶绿体是进行光合作用的场所。
- 1883年,德國的恩格尔曼運用三稜鏡將太陽光折射出各色光,照射絲狀海綿。一段時間後放入好氧細菌,發現植物在紅光與藍光區釋放較多氧氣
- 1897年,美国的Charles Reid Barnes首次在教科书中称它为光合作用。
- 1930年,C·B·凡尼尔藉由對紫硫菌的研究推論植物光合作用產生的氧來自水而非二氧化碳,革新了當時的觀念。
- 1941年,美国的鲁宾和卡门利用同位素标记法进行探究。证明光合作用释放氧气来自于水。
- 1945-1957年,梅尔文·卡尔文利用碳14當追蹤標的,找出植物將二氧化碳轉化成碳水化合物的途徑。
- 1961年,彼得·米切尔發表化學滲透理論解釋光反應中ATP的生成。
- 诺贝尔奖得主、科学家鲁道夫·马库斯发现电子转移链的作用和意义。
- 奥托·海因里希·瓦尔堡和迪安·伯克发现I-量子光合作用的反应,分裂二氧化碳,由呼吸作用激活。
- 2018年6月,美国《科学》杂志刊登的一项新研究说,蓝藻可利用近红外光进行光合作用,其机制与之前了解的光合作用不同。这一发现有望为寻找外星生命和改良作物带来新思路。新研究发现,上述蓝藻在有可见光的情况下,会正常利用"叶绿素a"进行光合作用,但如果处在阴暗环境中,缺少可见光,就会转为利用"叶绿素f",使用近红光进行光合作用。
原理
光合作用文字:二氧化碳+水+光能→葡萄糖+氧氣+水
植物與動物不同。綠色植物与部分微生物在陽光充足的白天,能利用太阳提供的能量來進行光合作用,以獲得生長發育必需的養分,这就是所谓的自營生物。此種自營又稱為光合自營。
這個過程的關鍵參與者是细胞内部的葉綠体。葉綠体在陽光的作用下,把經由氣孔進入葉片内部的二氧化碳和由根部吸收的水轉變成爲葡萄糖,同時釋放出氧氣:
- 12H2O + 6CO2 —hν→ (與葉綠素產生化學作用)C6H12O6 (葡萄糖) + 6O2 + 6H2O
:上式中等号两边的水不能抵消,虽然在化学上此方程式显得很特别。原因是左边的水分子是植物吸收所得,而且用于制造氧气和提供电子和氢离子。而右边的水分子的氧原子则是来自二氧化碳。为了更清楚地表达这一原料产物起始过程,人们更习惯在等号左右两边都写上水分子,或者在右边的水分子右上角打上星号。
植物的光合作用可分为光反应和碳反应两个步骤如下:
[光反應]

[碳反应]

光反应
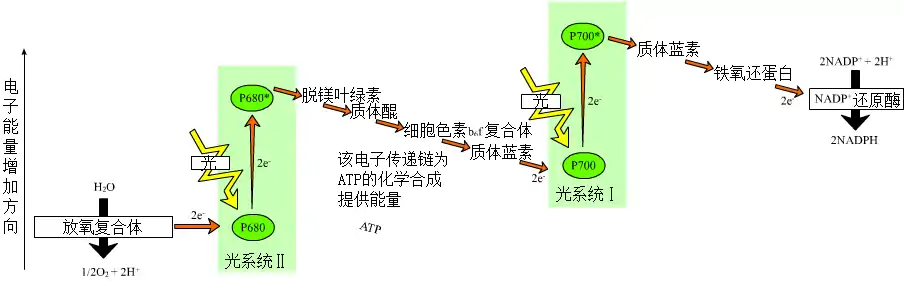
- 定义:光反应是反应中心色素所吸收的光能与原初电子受体和次级电子受体之间进行的氧化还原反应,从而实现光能转化为电能,并转变为化学能的过程。[5]
- 场所:类囊体膜
- 影响因素:光强度,水分供给,氧含量
- 过程:叶绿体膜上有两套光合作用系统(光系统):光合作用系统一和光合作用系统二(光合作用系统一比光合作用系统二要原始,但电子传递先在光合作用系统二开始,一、二的命名则是按其发现顺序)。在光照条件下,两系统分别吸收700nm和680nm波长的光子作为能量,将从水分子在光解过程中得到的电子不断传递,该过程还有细胞色素b6/f的参与。电子最后被传递给辅酶NADP,通过铁氧还蛋白-NADP还原酶将NADP还原为NADPH。而水光解所得的氢离子则顺浓度差通过类囊体膜上的蛋白质复合体而从类囊体内向外移动到基质,势能降低。其间的势能用于合成ATP,以供碳反应所用。此时势能已降低的氢离子则被氢载体NADP带走,一分子NADP可携带两个氢离子。这个NADPH與氢离子则在碳反应中充当还原剂的作用。
意义:
- 光解水,产生氧气。
- 将光能转变成化学能,产生三磷酸腺苷 (ATP),为碳反应提供能量。
- 利用水光解的产物氢离子,合成NADPH及氢离子,为碳反应提供还原剂。
詳細過程如下:
光系統由多種色素,如葉綠素a(Chlorophyll a)、葉綠素b(Chlorophyll b)、類胡蘿蔔素(Carotenoids)等組成。多种色素既拓寬了光合作用的作用光譜,其他的色素也能吸收過度的強光而產生所謂的光保護作用(Photoprotection)。在光系統裡,當光子击中系統裡的色素分子時,會如圖片(页面存档备份,存于) 所示,使電子在分子之間移轉,直到到达反應中心為止。反應中心有兩種,分别位于光系统一与光系统二。光系統一的吸收光譜於700nm達到峰值,系統二則以680nm為峰值。反應中心由葉綠素a及特定蛋白質所組成(這邊的葉綠素a是因為位置而非結構特殊),蛋白質的種類決定了反應中心吸收光線的波長。吸收相应波长的波長的光線後,葉綠素a激發出一個電子,而旁邊的酶使水光解成氫離子、电子和氧原子。此时多餘的電子去補葉綠素a分子上的缺少。然後葉綠素a透過如圖所示的過程,生產ATP與NADPH分子,该過程稱為電子傳遞鏈。
{kind=link}
{kind=link}
電子傳遞鏈分為兩種,即循環電子傳遞鏈(cyclic electron transfer system)和非循環電子傳遞鏈(noncyclic electron transfer system)
非循環電子傳遞鏈
非循環電子傳遞鏈過程大致如下:
從光系統2出發: 光系統2→初級接受者(Primary acceptor)→质体醌(Pq)→細胞色素複合體(Cytochrome Complex)→質體藍素(含銅蛋白質,Pc)→光系統1→初級接受者→鐵氧化還原蛋白(Fd)→NADP+還原酶(NADP+ reductase)
電子傳遞鏈從光系統2出發后,會光解水,釋放氧氣,產生三磷酸腺苷 (ATP)與NADPH。
循環電子傳遞鏈
循環電子傳遞鏈的過程如下:
從光系統1出發: 光系統1→初級接受者(Primary acceptor)→鐵氧化還原蛋白(Fd)→細胞色素複合體(Cytochrome Complex)→質體藍素(含銅蛋白的循環電子傳遞鏈不會產生氧氣,因為電子來源並非是水的光解),最後會生產出三磷酸腺苷(ATP)。
在非循環電子傳遞鏈中,細胞色素複合體會將氫離子打到類囊體(Thylakoid)裡面。高濃度的氫離子會順浓度差向類囊體外擴散。由于類囊體膜由雙層磷脂分子组成,所以氢离子必須靠通道蛋白進行擴散,這個通道蛋白就是ATP合酶。
固碳作用(碳反应)
固碳作用实质上是一系列酶促反应的结果。生物界有几种固碳方法,其中以卡尔文循环为主,但并非所有进行光合作用的细胞都使用卡尔文循环进行碳固定,例如绿硫细菌会使用还原性三羧酸循环,绿曲挠菌(Chloroflexus)会使用3-羟基丙酸途径(3-Hydroxy-Propionate pathway),还有一些生物会使用核酮糖-单磷酸途径(Ribolose-Monophosphate Pathway)和丝氨酸途径(Serine Pathway)进行碳固定。
- 场所:叶绿体基质
- 影响因素:温度,二氧化碳浓度
- 过程:不同的植物,固碳作用的过程不一样,而且叶片的解剖结构也不相同。这是植物对环境的适应的结果。固碳作用可分为C3,C4和CAM(景天酸代谢)三种类型(见下文)。三种类型由二氧化碳固定过程的不同而划分。
卡爾文循環
卡尔文循环是光合作用裡碳反应的一部分。反应场所为叶绿体内的基质。循环可分为三个阶段:羧化、还原和二磷酸核酮糖的再生。大部分植物会将吸收到的一分子二氧化碳,通过核酮糖-1,5-二磷酸羧化酶/加氧酶的作用,整合到一个五碳糖分子1,5-二磷酸核酮糖(RuBP)的第二位碳原子上。此过程称为二氧化碳的固定。这一步反应的意义是,把原本并不活泼的二氧化碳分子活化,使之随后能被还原。但这种六碳化合物極不稳定,会分解为两分子的三碳化合物3-磷酸甘油酸。后者被在光反应中生成的NADPH与氢离子还原,此过程需要消耗ATP。产物是3-磷酸丙糖。后来经过一系列复杂的生化反应,一个碳原子将会被用于合成葡萄糖而离开循环。剩下的五个碳原子经一系列变化,最后再生成一分子1,5-二磷酸核酮糖,循环重新开始。卡尔文循环每进行六次,生成一分子葡萄糖。
各类生物的光合作用
C3类植物
二次大战后,美国加州大学柏克萊分校的梅尔文·卡尔文与他的同事们研究一种綠球藻,以确定植物在光合作用中如何固定CO2。此时C14示踪技术和双向纸层析法技术都已成熟,卡尔文正好在实验中用上此两种技术。
他们将培养出来的綠球藻,放置在含有未标记CO2的密闭容器中,然后将C14标记的CO2注入容器,培养相当短时间后,将綠球藻浸入热的乙醇中杀死细胞,使细胞中的酶变性而失效。接着他们提取到溶液裡的分子。然后将提取物应用双向纸层析法,分离各种化合物,再通过放射自显影,分析放射性上面的斑点,并与已知化学成份比较。
卡尔文在实验中发现,标记有C14的CO2很快就能转变成有机物。在几秒钟内,层析纸上就出现放射性斑点,经与已知化学物比较,斑点中的化学成分是3-磷酸甘油酸(3-phosphoglycerate,PGA),是糖酵解的中间体。这第一个被提取到的产物是一个三碳分子,所以将这种CO2固定途径称为C3途径,将通过这种途径固定CO2的植物称为C3植物。后来研究还发现,CO2固定的C3途径是一个循环过程。人们称之为C3循环。这一循环又称「卡尔文循环」。
C3类植物,如米和麦,二氧化碳经气孔进入叶片后,直接进入叶肉进行卡尔文循环。而C3植物的维管束鞘细胞 (bundle-sheath cells) 很小,不含或含很少叶绿体,卡尔文循环不在这里发生。
C4类植物
1960年代,澳洲科学家哈奇(M. D. Hatch)和斯莱克(C. R. Slack)发现玉米、甘蔗等热带绿色植物,除了和其他绿色植物一样,具有卡尔文循环外,CO2首先通过一条特别的途径被固定。这条途径也被称为「哈奇-斯莱克途径」。
C4植物主要是一些生活在干旱热带地区的植物。在这种环境中,植物倘若长时间开放气孔吸收二氧化碳,会导致水分通过蒸腾作用过快的流失。所以,植物只能短时间开放气孔,二氧化碳摄入量必然少。植物必须利用这少量的二氧化碳进行光合作用,合成自身所需物质。
在C4植物叶片维管束的周围,有维管束鞘围绕,这些维管束鞘细胞含有叶绿体,但裡面并无基粒或发育不良。在这裡,就是主要进行卡尔文循环的地方。
其叶肉细胞中,含有独特的酶,即磷酸烯醇式丙酮酸羧基化酶,使得二氧化碳先被一种三碳化合物磷酸烯醇式丙酮酸同化,形成四碳化合物草酰乙酸,这也是该暗反应类型名称的由来。这草酰乙酸在转变为苹果酸盐后,进入维管束鞘,就会分解释放二氧化碳和一分子丙酮酸。二氧化碳利用此细胞才进入卡尔文循环,后同C3进程。而丙酮酸则会被再次合成磷酸烯醇式丙酮酸。此过程消耗ATP。
该类型的优点是,二氧化碳固定效率比C3高很多,有利植物在干旱环境生长。C3植物行光合作用所得的淀粉,会贮存在叶肉细胞中,因为这是卡尔文循环的场所,而维管束鞘细胞则不含叶绿体。而C4植物的淀粉,将会贮存于维管束鞘细胞 (bundle-sheath cells) 内,因为C4植物的卡尔文循环是在此发生的。
景天酸代謝(CAM)植物
如果说C4植物是空间上错开二氧化碳的固定和卡尔文循环的话,那景天酸循環就是时间上错开这两者。行使这一途径的植物,是那些有着膨大肉质叶子的植物,如凤梨。这些植物晚上开放气孔,吸收二氧化碳,同样经哈奇-斯莱克途径将CO2固定。早上的时候气孔关闭,避免水分流失过快。同时在叶肉细胞中進行卡尔文循环。这些植物二氧化碳的固定效率也很高。
意义:二氧化碳的固定,使得原本化学性质不活泼的二氧化碳,化学活性增加,以利于被还原,最后合成葡萄糖。
参考文献
- 沈允钢. . 生物學文摘. 2006, 20 (2): 1–1 [2020-08-01]. (原始内容存档于2020-08-18). 已忽略未知参数
|注=(帮助) - Miyamoto K. . Renewable biological systems for alternative sustainable energy production (FAO Agricultural Services Bulletin - 128). Food and Agriculture Organization of the United Nations. [2009-01-04]. (原始内容存档于2013-09-07).
- Zhu, Xin-Guang; Long, Stephen P.; Ort, Donald R. . Annual Review of Plant Biology. 2010-06-02, 61 (1). ISSN 1543-5008. doi:10.1146/annurev-arplant-042809-112206 (英语).
- https://zhuanlan.zhihu.com/p/22734035
- 《植物生物学》第四版 周云龙 刘全儒