ABO血型系统
ABO血型系统是人类最早认识也是最为重要的血型系统。ABO血型由红细胞膜上的不同抗原所决定,与人类输血时发生的溶血反应密切相关,具有重要的临床意义。ABO抗原也存在于牛、羊和一些猿类等其他动物体内[1]。ABO血型可能繼承自數百萬年前一個共同的靈長類祖先,並在傳染病免疫反應具關鍵作用[2]。
发现史
1900年,奥地利维也纳大学病理研究所的生物学家卡尔·兰德施泰纳首次报道,健康人的血清对一些人类个体的红细胞有凝聚作用。通过混合不同人的血清和红细胞,他发现了A、B、O三种血型[3],他的学生和又于两年后发现了第四种——AB型[4]。兰德施泰纳因此获得1930年度诺贝尔生理学或医学奖。捷克血清学家扬·扬斯基也于1907年独立发现人类血液可分为四种血型[5]。但由于当时通讯不便,只有兰德施泰纳的成就得到科学界广泛承认,扬斯基则几乎被遗忘,仅在前苏联等少数国家仍有提及。
1910至1911年,波兰人希尔斯菲尔德和德国人埃米爾·馮·登格恩发现了ABO血型系统的遗传性。1924年,德国数学家费利克斯·伯恩斯坦进一步阐释了ABO的遗传方式是由同一基因座上的几个等位基因决定的[6]。英国人沃特金斯和摩尔根发现,决定ABO抗原不同的分子基团(称为抗原决定簇或表位,)由糖类转化而来,决定A抗原的是N-乙酰半乳糖胺(N-acetylgalactosamine),决定B抗原的是半乳糖[7]。1990年,日本学者山本等首次描述了编码这些抗原决定簇的A、B、O等位基因的DNA结构[8]。
ABO抗原
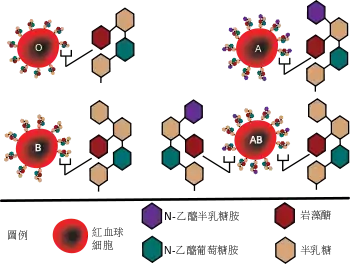
人类ABO血型系统的抗原合成基于前体物质H抗原。H抗原是一种糖脂,基本分子结构是以糖苷键与多肽链骨架结合的四糖链,即β-D-半乳糖、β-D-N-乙酰葡萄糖胺、β-D-半乳糖以及在β-D-半乳糖2-位连接的抗原决定簇α-L-岩藻糖。H抗原的决定基因FUT1位于19号染色体,长度超过5000个碱基对,包括3个外显子。FUT1基因有两个等位基因H和h,H等位基因编码岩藻糖转移酶,使岩藻糖与糖链末端的半乳糖相连,形成H抗原。而h等位基因无法编码具有活性的岩藻糖转移酶。hh纯合子个体在人类中非常罕见,形成所谓孟买血型。参见Hh/孟买血型系统。
编码ABO抗原决定簇的是9号染色体上的ABO基因,总长度18000至20000个碱基对,包括7个外显子,其中最大的第7外显子和第6外显子的碱基数占整个编码序列的77%[9]。ABO基因有三个最主要的等位基因:IA(A)、IB(B)和i(O),这些等位基因的原初产物是糖基转移酶。IA等位基因编码α-1,3N-乙酰氨基半乳糖转移酶,能将α-N-乙酰半乳糖胺接到H抗原的β-D-半乳糖上,形成A抗原;IB等位基因编码α-1,3-D-半乳糖转移酶,将α-D-半乳糖接到H抗原的相同位置,形成B抗原;i等位基因的第6外显子包含一个核苷酸缺失,导致其编码的蛋白质无法正常表达,从而失去酶活性,因此,O型血的抗原就是未经改变的H抗原。
A、B抗原主要表达在红细胞膜上,以糖蛋白或糖脂的形式,呈树枝状突出细胞表面。在大多数上皮细胞、内皮细胞以及体液中,也有A、B抗原的存在。
亚型
1930年,人们发现A抗原可以分为A1与A2两个亚型。目前已知,A型至少包括20个亚型,绝大多数是A1和A2(>99%),其中A1的比例超过80%[10]。 1980年以前,人们通常认为红细胞的ABH决定簇主要存在于鞘糖脂分子上,但是,1980年发表的几篇报道指出:糖脂与糖蛋白相比,在ABH抗原决定簇中仅扮演次要角色。红细胞上的多数ABH抗原都位于离子交换蛋、带3蛋白、葡萄糖转运蛋白以及带4.5蛋白的单个高度分枝的多聚N-乙酰氨基乳糖的N-型糖苷键上。
血清学

根据红细胞膜上A抗原与B抗原的有无,可以将血液分为4型:如果只存在A抗原,则称为A型;只存在B抗原,则称为B型;若A与B两种抗原都存在,则称为AB型;这两种抗原都没有的,则称为O型。
不同血型的血液里可能存在抗-A或抗-B抗体(也称血凝集素,见右图)。这些抗体通常是免疫球蛋白IgM,无法通过胎盘从母体进入胎儿血液,但会在出生后一年内出现在人体内,产生原因可能是环境中某些与A、B抗原决定簇类似的糖类的刺激。这些抗体遇到与本身血型不合的红细胞时,就会产生溶血反应,例如一个A血型的人如果输入B型血,会立刻导致体内的抗-B抗体与B型红细胞的表面B抗原结合,引发补体介导的红细胞自溶。相同的抗原-抗体反应也会在B血型的人输入A型血或O型血的人输入其他血型的血液时发生。只有AB血型的人体内没有抗A和抗B,可以接受其他血型的血液,故曾被称为“万能受血者”;而各种血型的人均可以输入O型血,因此O型血的人曾被称为“万能供血者”(见下表)。然而,这种“万能”的说法并不严格。以A血型的受血者输入O型血为例,O型血的红细胞不含B抗原,不会与受血者体内的抗-B反应,但O型血中的抗-A会与受血者血液中带有A抗原的红细胞反应。当输血量较小时,输入的抗-A抗体被受血者的血清充分稀释,其影响可以降低;但如果输血量较大,抗-A无法得到充分稀释,仍可能引发受血者的溶血反应。在這種情況下,一個解決之道是將送血者血液中的抗體分離,否則不应进行异型输血。
| 受血者 | 供血者 |
|---|---|
| A型 | A或O型 |
| B型 | B或O型 |
| AB型 | A、B、AB或O型 |
| O型 | O型 |
遗传性
ABO血型系统的遗传是单基因决定的。ABO基因位于9号染色体的长臂上(9q34),有三个主要的等位基因IA(A)、IB(B)和i(O)。IA和IB的产物是具有活性的酶,分别催化A抗原和B抗原的合成,而等位基因i的产物不具有酶活性,无法催化A、B抗原合成。由于人类染色体是双倍体,一个人通常只能拥有三个等位基因中的两个,分别来自父母双方。这两个等位基因的类型,即血型的基因型(genotype),决定了人类血型的表现型(phenotype)。
IA和IB对i均为显性,故而只有基因型是ii的人才有O型血,基因型是IAIA或IAi的人是A血型,基因型是IBIB或IBi的人是B血型。而IA和IB是共显性,因此基因型是IAIB的人具有两种表型,即AB血型。
一般说来,如果父母双方均为O型血,子女必然是O型;如果父母有一方是AB型血,子女不可能是O型;A型和O型血的父母不可能生育B型或AB型血的子女,B型和O型血的父母不可能生育A型或AB型血的子女。因此,ABO血型系统曾经广泛用于亲子鉴定。父母与子女血型的详细关系及子女血型的可能性见下表:
| 父母血型/基因型 | A型 | B型 | O型(OO) | AB型(AB) | |||
|---|---|---|---|---|---|---|---|
| AA | AO | BB | BO | ||||
| A型 | AA | A型(AA) 100% | A型 100% (AA 50%、AO 50%) |
AB型 100% | A型(AO) 50% AB型 50% |
A型(AO) 100% | A型(AA) 50% AB型 50% |
| AO | A型 100% (AA 50%、AO 50%) |
A型 75% (AA 25%、AO 50%) O型 25% |
B型(BO) 50% AB型 50% |
A型(AO) 25% O型 25% B型(BO) 25% AB型 25% |
A型(AO) 50% O型 50% |
A型 50% (AA 25%、AO 25%) B型(BO) 25% AB型 25% | |
| B型 | BB | AB型 100% | B型(BO) 50% AB型 50% |
B型(BB) 100% | B型 100% (BB 50%、BO 50%) |
B型(BO) 100% | B型(BB) 50% AB型 50% |
| BO | A型(AO) 50% AB型 50% |
A型(AO) 25% O型 25% B型(BO) 25% AB型 25% |
B型 100% (BB 50%、BO 50%) |
B型 75% (BB 25%、BO 50%) O型 25% |
B型(BO) 50%、 O型 50% |
B型 50% (BB 25%、BO 25%) A型(AO) 25% AB型 25% | |
| O型(OO) | A型(AO) 100% | A型(AO) 50% O型 50% |
B型(BO) 100% | B型(BO) 50% O型 50% |
O型 100% | A型(AO) 50% B型(BO) 50% | |
| AB型(AB) | A型(AA) 50% AB型 50% |
A型 50% (AA 25%、AO 25%) B型(BO) 25% AB型 25% |
B型(BB) 50% AB型 50% |
B型 50% (BB 25%、BO 25%) A型(AO) 25% AB型 25% |
A型(AO) 50% B型(BO) 50% |
A型(AA) 25% B型(BB) 25% AB型 50% | |
cis-AB
极少数人类个体的一条染色体中同时存在一个cis-AB基因,它编码的酶可以同时催化A抗原和B抗原的合成。cis-AB是从一个亲本遗传而来的,因此即便其基因型是cisAB / O,也会表现为AB型,若与O型血的人(基因型O / O)生育,子女的血型可能是AB或O,而不像通常AB型血与O型血的父母只能有A或B型血的子女。
ABO血型在人群中的分布
随着地域、人群的不同,ABO各个血型的频率也有很大区别,常作为研究人类的民族分化、迁徙史的工具。
世界人群
从世界角度来看,IB等位基因的频率最低,约为16%,在中亚一带的阿爾泰民族最常见,而在美洲和澳大利亚原住民中最为罕见。IA等位基因较IB更为常见,约21%的世界人口携带IA,但相对高频率的区域比较分散,如北美印第安人黑脚族(30-35%),澳大利亚一些原住民族群(40-53%),斯堪的納維亞地區北部的薩米人(50-90%)等,而在中南美的印第安人中,IA几乎无分布。由于IA、IB两种等位基因型频率不高,O血型称为世界上最常見的血型,平均频率达63%,特别是在中南美洲印第安人中,O型血频率接近100%,O血型在澳大利亚原住民和西歐人群(特别是凯尔特人)中也很常见,在東歐和中亚的频率则相对较低。[11] A型在北欧西欧与日本、湖南最多。
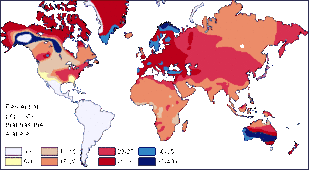 A型分布
A型分布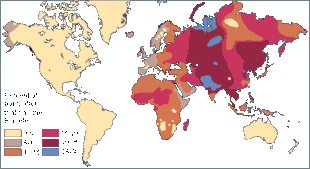 B型分布
B型分布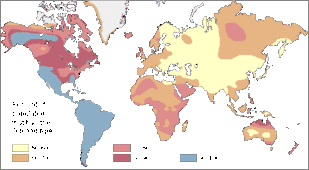 O型分布
O型分布
參見
- 血型性格學說
参考文献
- Maton, Anthea; Jean Hopkins; Charles WilliamMcLaughlin; Susan Johnson; Maryanna Quon Warner; David LaHart; Jill D. Wright. . Englewood Cliffs, New Jersey, USA: Prentice Hall. 1993. ISBN 0-13-981176-1.
- . [2012-10-27]. (原始内容存档于2012-10-25).
- Landsteiner, Karl. . Zentralblatt Bakteriologie. 1900 [2022-08-07]. OCLC 78393656. (原始内容存档于2022-08-07) (德语).
- Pullini, T. . Archiv der Pharmazie. 1836, 57 (2) [2022-08-07]. ISSN 0365-6233. doi:10.1002/ardp.18360570225. (原始内容存档于2022-08-07) (德语).
- Janský J. . Sborn. Klinick. 1907, 8: 85–139 (捷克语).
- Crow, J F. . Genetics. 1993-01-01, 133 (1) [2022-08-07]. ISSN 1943-2631. PMC 1205297
 . PMID 8417988. doi:10.1093/genetics/133.1.4. (原始内容存档于2022-08-07) (英语).
. PMID 8417988. doi:10.1093/genetics/133.1.4. (原始内容存档于2022-08-07) (英语). - Morgan, W. T. J. & Watkins, W. M. Br. Med. Bull. 25, 30–34 (1969), Watkins, W. M. in: Advances in Human Genetics Vol. 10 (eds Harris, H. & Hirschhorn, K.) 1–136 (Plenum, New York, 1980), Watkins, W. M. & Morgan, W. T. J. Vox Sang. 4, 97−119 (1959)
- Yamamoto, Fumi-ichiro; Clausen, Henrik; White, Thayer; Marken, John; Hakomori, Sen-itiroh. . Nature. 1990-05, 345 (6272) [2022-08-07]. ISSN 1476-4687. doi:10.1038/345229a0. (原始内容存档于2022-08-07) (英语).
- Olsson ML, Chester MA. . Transfution Med. 2001, 11: 295–313.
- Blood Group A Suptypes (页面存档备份,存于),The Owen Foundation, retrieved 2008-07-01.
- . [2008-12-25]. (原始内容存档于2009-12-11).