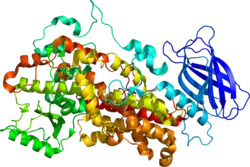



ALOX12 (EC 1.13.11.31), also known as arachidonate 12-lipoxygenase, 12-lipoxygenase, 12S-Lipoxygenase, 12-LOX, and 12S-LOX is a lipoxygenase-type enzyme that in humans is encoded by the ALOX12 gene which is located along with other lipoyxgenases on chromosome 17p13.3.[5][6] ALOX12 is 75 kilodalton protein composed of 663 amino acids.
Nomenclature
| arachidonate 12-lipoxygenase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| EC no. | 1.13.11.31 | ||||||||
| CAS no. | 82391-43-3 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| Gene Ontology | AmiGO / QuickGO | ||||||||
| |||||||||
Other systematic names for ALOX12 include 12S-Lipoxygenase, platelet-type 12-lipoxygenase, arachidonate:oxygen 12-oxidoreductase, Delta12-lipoxygenase, 12Delta-lipoxygenase, and C-12 lipoxygenase. ALOX12, often termed plate platelet-type 12-lipoxygenase, is distinguished from leukocyte-type 12-lipoxygenase which is found in mice, rats, cows, and pigs but not humans. Leukocyte-type 12-lipoxygenase in these animal species shares 73-86% amino acid identity with human ALOX15 but only 57-66% identity with human platelet-type 12-lipoxygenase and, like ALOX15, metabolizes arachidonic acid primarily to 15(S)-hydroperoxy-5Z,8Z,11Z,13E-eicosatetraenoic acid (i.e. 15(S)-HpETE; see 15-Hydroxyeicosatetraenoic acid).[7] Accordingly, rodent leukocyte 12-lipoxygenase is deemed an ortholog of ALOX15 and is designated as Alox15.[8]
Human ALOX12 and ALOX15 along with rodent leukocyte-type Alox12 and Alox15 are commonly termed 12/15-lipoxygenases based on their ability to metabolize arachidonic acid to both 12(S)-HpETE and 15(S)-HpETE and to conduct this same metabolism on arachidonic acid that is esterified to membrane phospholipids; human ALOX15B makes 15(S)-HpETE but not 12(S)-HpETE and therefore is not regarded as a 12/15-lipoxygenase.[9] Studies on the role of ALOX12 in pathophysiology using the main models for such functional studies, rats and mice, are complicated because neither species possesses a lipoxygenase that makes a predominance of 12(S)-HETE and therefore is metabolically equivalent to ALOX12.[7][9] For example, the functions inferred for Alox12 in mice made deficient in Alox12 using knockout methods may not indicate a similar function for ALOX12 in humans due to differences in these two enzymes' metabolic activities. The function of ALOX12 is further clouded by human ALOX15 which metabolizes arachidonic acid primarily to 15(S)-HpETE but also makes lesser but still significant amounts of 12(S)-HpETE (see ALOX15).
ALOX12 is also distinguished from arachidonate 12-lipoxygenase, 12R type (ALOX12B), which metabolizes arachidonic acid to the R stereoisomer of 12(S)-HpETE viz., 12(R)-hydroperoxy-5Z,8Z,10E,14Z-icosatetraenoic acid (12(R)-HpETE), a product with very different pathophysiological roles than that of 12(S)-HpETE (see ALOX12B).
Discovery
ALOX12, originally called arachidonate 12-lipoxygenase, was first characterized by the Nobel Laureate, Bengt I. Samuelsson, and his famed colleague, Mats Hamberg, in 1974 by showing that human platelets metabolize arachidonic acid not only by the well-known cyclooxygenase pathway into prostaglandins and 12-hydroxyheptadecatrienoic acid but also by a cyclooxygenase-independent pathway to 12(S)-hydroperoxy-5,8,10,14-eicosatetraenoic acid; this activity was the first mammalian lipoxygenase activity to be characterized.[10] In 1975, the first biological activity was attached to this metabolite in studies showing that it simulated the chemotaxis of human neutrophils.[11] During the several years thereafter, human ALOX12 was purified, characterized biochemically, and had its gene molecularly cloned.[7][12]
Tissue distribution
Based predominantly on the presence of its mRNA, human ALOX12 is distributed predominantly in blood platelets and leukocytes and at lower levels in the basal layer of the epidermis (particularly in the skin lesions of psoriasis), islets of Langerhans within the pancreas, and certain cancers.[13]
Enzyme activities
The control of ALOX12 activity appears to rest principally on the availability of its polyunsaturated fatty acid (PUFA) substrates which are released from storage in membrane phospholipids by cell stimulation.[14] The enzyme participates in arachidonic acid metabolism by conducting the following chemical reaction wherein its substrates are arachidonic acid (also termed as arachidonate or, chemically, as 5Z,8Z,11Z,14Z-eicosatetraenoic acid) and O2 (i.e. oxygen) and its product is 12S-hydroperoxy-5Z,8Z,10E,14Z-eicosatetraenoic acid (i.e. 12S-hydroperoxyeicosatetraenoic acid or 12S-HpETE):[10][15]
- arachidonate + O2 → 12S-hydroperoxy-5Z,8Z,10E,14Z-eicosatetraenoic acid
In cells, 12SHpETE may be further metabolized by ALOX12 itself, by ALOXE3 or possibly other, as yet not fully identified, hepoxilin syntheses to hepoxilin A3 (8R/S-hydroxy-11,12-oxido-5Z,9E,14Z-eicosatrienoic acid) and B3 (10R/S-hydroxy-11,12-oxido-5Z,8Z,14Z-eicosatrienoic acid):[16][17][18]
- 12S-hydroperoxy-5Z,8Z,10E,14Z-eicosatetraenoic acid → 8R/S-hydroxy-11,12-oxido-5Z,9E,14Z-eicosatrienoic acid + 10R/S-hydroxy-11,12-oxido-5Z,8Z,14Z-eicosatrienoic acid
Hepoxilins can promote certain inflammation responses, increase pain perception (i.e. tactile allodynia), regulate regional blood flow, and contribute to the regulation of blood pressure in animal models (see Hepoxilins). Far more commonly, however, 12S-HpETE is rapidly reduced to its hydroxyl product by ubiquitous cellular peroxidase activities thereby forming 12S-hydroxy-5Z,8Z,10E,14Z-eicosatetraenoic acid, i.e. 12-hydroxyeicosatetraenoic acid or 12S-HETE:[19]
- 12S-hydroperoxy-5(Z),8(Z),10(E),14(Z)-eicosatetraenoic acid → 12S-hydroxy-5(Z),8(Z),10(E),14(Z)-eicosatetraenoic acid
12S-HETE promotes inflammation responses, may be involved in the perception of puritis (i.e. itching) in the skin, and regulates regional blood flow in animal models; it also promotes the malignant behavior of cultured human cancer cells as well as the growth of certain cancers in animal models (see 12-HETE). While arachidonate and 12(S)-HETE are the predominant substrates and products, respectively, of ALOX12, the enzyme also metabolizes other PUFA.
It metabolizes the omega-3 fatty acid, docosahexaenoic acid (DHA i.e., 4(Z),7(Z),10(Z),13(Z),16(Z),19(Z)-docosahexaenoic acid to 14(R)-hydroperoxy-4(Z),8(Z),10(Z),12(E),16(Z),19(Z)-docosahexaenoic acid)(i.e. 17-hydroperoxy-DHA)
Then, ALOX12 or an unidentified epoxidase-type enzyme may metabolize this intermediate to an epoxide, 13,14-epoxy-4(Z),7(Z),9(E),11(E),16(Z),19(Z)-docosahexaenoic acid (i.e. 13,14-e-maresin)
This is further metabolized to 7R,14S-dihydroxy-4Z,8E,10E,12Z,16Z,19Z-docosahexaenoic acid (i.e. Maresin 1), by an unidentified epoxide hydrolase-type enzyme:
- DHA → 17-hydroperoxy-DHA → 13,14-e-maresin → Maresin-1
Maresin 1 has a set of activities that may oppose those of 12(S)-HETE and the hepoxilins; it is a member of a class of PUFA metabolites termed Specialized pro-resolution mediators (SPMs) which possess anti-inflammatory, pain-alleviating, and other defensive activities.[20] ALOX12 also acts on leukotriene A4 (LTA4) in a two cellular reaction termed transcellular metabolism: human neutrophils metabolize arachidonic acid to its 5,6-epoxide, LTA4, and releases this intermediate to nearby neutrophils which metabolize it to lipoxin A4 (5S,6R,15S-trihydroxy-7E,9E,11Z,13Z-eicosatetraenoic acid) and lipoxin B4 (5S,14R,15S-trihydroxy-6E,8Z,10E,12E-eicosatetraenoic acid); both lipoxins are SPMs with many SPM-like activities (see lipoxin).[21] ALOX12 may also metabolize lesser amounts of DHA to secondary products including 17-hydroperoxy-DHA, 11-hydroperoxy-DHA, and 8,14-dihydroxy-DHA[20] ALOX12 may likewise metabolize 5(S)-HETE to 5S,12S-dihydroxyeicosatetraenoic acid (12,15-diHETE) and 15S-HETE to 14,15S-diETE.[14] While these compounds have not been thoroughly evaluated for bioactivity, 17-hydroperoxy-HDHA and the reduced product to which it is rapidly converted in cells, 17-hydroxy-HDHA, have been shown to inhibit the growth of cultured human prostate cancer cell by causing them to enter apoptosis.[22]
Animal studies
Studies on rodents lacking or made deficient in the leukocyte-type 12-lipoxygenase, Alox12 (which is most closely related to human ALOX15) implicate this enzyme in: a) preventing the development and complications of dietary-induced and/or genetically induced diabetes, adipose cell/tissue dysfunction, and obesity; b) the development of atherosclerosis and Steatohepatitis; b) regulating blood vessel contraction, dilation, pressure, remodeling, and angiogenesis; c) maintaining normal renal, neurological, and brain function; and d) the development of Alzheimer's disease.[8][9][23] In these studies, it is usually unclear which, if any metabolite(s) of Alox12 was implicated.
Preclinical studies
Metabolic syndrome
The metabolic syndrome is a clustering of at least three of five of the following medical conditions: abdominal (central) obesity, elevated blood pressure, elevated fasting plasma glucose (or overt diabetes), high serum triglycerides, and low high-density lipoprotein (HDL) levels. ALOX12 and its metabolite, 12(S)-HETE, are elevated in the islets of Langerhans of patients with type 1 diabetes or type 2 diabetes as well as in the fat cells of white adipose tissue of morbidly obese type 2 diabetic patients.[8] The PP cells (i.e. gamma cells) of the pancreas islets appear to be the major if not only site where ALOX12 is expressed in these patients.[8] The studies propose that in the islets of Langerhans ALOX12 and its 12(S)-HETE product cause excessive production of reactive oxygen species and inflammation which lead to losses in insulin-secreting beta cells and thereby types 1 and 2 diabetes and that in adipose tissue the excess in AlOX12, 12(S)-HETE, reactive oxygen species, and inflammation lead to fat cell dysfunction (also see 12-HETE#Inflammation and inflammatory diseases and 12-HETE#Diabetes). Indeed, in one study a Single-nucleotide polymorphism, rs2073438,[24] located in an intron region of the ALOX12 gene was significantly associated with total and percentage fat mass of obese compared to non-obese young Chinese men.[8][13][18] ALOX12 and 12(S)-HETE are likewise implicated in essential hypertension (see next section). Hence, ALOX12 and its metabolite(s) may contribute to the development and/or progression of obesity, diabetes, hypertension, and/or the metabolic syndrome.
Blood vessels
A selective but not totally specific inhibitor of ALOX12 reduced the growth response of cultured human endothelial cells to basic fibroblast growth factor and vascular endothelial growth factor (VEGF); this reduction was partially reversed by 12(S)-HETE; 12(S)-HETE also stimulates human prostate cell lines to produce VEGF.[19] These results suggest that growth responses to the two growth factors involves their stimulation of 12(S)-HETE production by endothelial cells and therefore that ALOX12 may be a target for reducing the neo-vascularization that promotes arthritic and cancer diseases. 12(S)-HETE also dilates human coronary microcirculation arteries by activating these vessels' smooth muscle BKca Potassium channels and is therefore suggested to be an Endothelium-derived hyperpolarizing factor.[9][19] Finally, a single nucleotide variant in the ALOX12 gene (R261Q [3957 G>A]) has been associated with essential hypertension and elevation in the urinary excretion of 12(S)-HETE in humans and may be a contributing factor for to essential hypertension (see also 12-HETE#Blood pressure).[9][25]
Alzheimer's disease
Patients with Alzheimer's disease or other forms of dementia have significantly higher levels of 12(S)-HETE (and 15(S)-HETE) in cerebrospinal fluid compared to aged-matched normal individuals. Complementary studies in rodent models bearing human mutated genes for Amyloid precursor protein and/or tau protein (see tau protein#Clinical significance) that produce Alzheimer's dementia-like syndromes implicate 12(S)-HETE, 15(S)-HETE, and a 12/15-lipoxygenase type enzyme in the development and progression of the Alzhiemer's disease-like symptoms and findings in these animals.[23] In a single study, ALOX12 mRNA was found elevated in the brain tissue of Alzheimer disease patients compared to control patients.[13] These results suggest that ALOX12 (or ALOX15) may contribute to the development of Alzheimer's disease in humans.
Cancer
Studies in prostate cancer find that human prostate cancer cell lines in culture overexpress ALOX12, overproduce 12(S)-HETE, and respond to 12(S)-HETE by increasing their rate of proliferation, increasing their cell surface expression of integrins, increasing their survival and delaying their apoptosis, and increasing their production of vascular endothelial growth factor and MMP9 (i.e. Matrix metallopeptidase 9); selective (but not entirely) specific ALOX12 inhibitors reduced the proliferation and survival of these cells (see also 12-HETE#prostate cancer). These finding suggest that ALOX12 and its 12(S)-HETE product may contribute to the growth and spread of prostate cancer in humans.[19] Recently, hypermethylation of the ALOX12 gene in prostate cancer tissue was associated with clinical predictors for a high rate of recurrent disease.[26] Some studies have found that 12(S)-HETE also promotes the growth and/or related pro-malignant behaviors of various other types of cultured cancer cell lines (see 12-HETE#Other cancers).[19] ALOX12 has been shown to interact with Keratin 5 and LMNA as screened in a yeast two-hybrid interaction library from human epidermoid carcinoma A431 cells; these proteins are candidates for regulating 12-LOX, particularly in tumor cells.[27]
Platelet function
Although first identified in human platelets, the role of ALOX12 and its major metabolites, 12(S)-HpETE and 12(S)-HETE in platelet function remains controversial and unclear; it is possible that the ALOX12-12(S)-HETE metabolic pathway has dual functions in promoting or inhibiting platelet responses depending on the stimulating agent and response studied but that inhibiting ALOX12 may ultimately prove useful in inhibiting platelet-related blood clotting.[19]
Other associations
The ALOX12 gene has susceptibility alleles (rs6502997,[28] rs312462,[29] rs6502998,[30] and rs434473[31]) for the parasitic disease, human congenital toxoplasmosis.[13][32] Fetus bearer of these alleles thus suffer an increased susceptibility to this disease.
References
- 1 2 3 GRCh38: Ensembl release 89: ENSG00000108839 - Ensembl, May 2017
- 1 2 3 GRCm38: Ensembl release 89: ENSMUSG00000000320 - Ensembl, May 2017
- ↑ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ↑ "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ↑ Funk CD, Funk LB, FitzGerald GA, Samuelsson B (May 1992). "Characterization of human 12-lipoxygenase genes". Proceedings of the National Academy of Sciences of the United States of America. 89 (9): 3962–6. Bibcode:1992PNAS...89.3962F. doi:10.1073/pnas.89.9.3962. PMC 525611. PMID 1570320.
- ↑ "Entrez Gene: ALOX12 arachidonate 12-lipoxygenase".
- 1 2 3 Yamamoto S, Suzuki H, Ueda N (March 1997). "Arachidonate 12-lipoxygenases". Progress in Lipid Research. 36 (1): 23–41. doi:10.1016/s0163-7827(97)00002-7. PMID 9373619.
- 1 2 3 4 5 Tersey SA, Bolanis E, Holman TR, Maloney DJ, Nadler JL, Mirmira RG (June 2015). "Minireview: 12-Lipoxygenase and Islet β-Cell Dysfunction in Diabetes". Molecular Endocrinology. 29 (6): 791–800. doi:10.1210/me.2015-1041. PMC 4447641. PMID 25803446.
- 1 2 3 4 5 Dobrian AD, Lieb DC, Cole BK, Taylor-Fishwick DA, Chakrabarti SK, Nadler JL (January 2011). "Functional and pathological roles of the 12- and 15-lipoxygenases". Progress in Lipid Research. 50 (1): 115–31. doi:10.1016/j.plipres.2010.10.005. PMC 3012140. PMID 20970452.
- 1 2 Hamberg M, Samuelsson B (September 1974). "Prostaglandin endoperoxides. Novel transformations of arachidonic acid in human platelets". Proceedings of the National Academy of Sciences of the United States of America. 71 (9): 3400–4. Bibcode:1974PNAS...71.3400H. doi:10.1073/pnas.71.9.3400. PMC 433780. PMID 4215079.
- ↑ Turner SR, Campbell JA, Lynn WS (June 1975). "Polymorphonulcear leukocyte chemotaxis toward oxidized lipid components of cell membranes". The Journal of Experimental Medicine. 141 (6): 1437–41. doi:10.1084/jem.141.6.1437. PMC 2189855. PMID 1127383.
- ↑ Yoshimoto T, Yamamoto Y, Arakawa T, Suzuki H, Yamamoto S, Yokoyama C, Tanabe T, Toh H (November 1990). "Molecular cloning and expression of human arachidonate 12-lipoxygenase". Biochemical and Biophysical Research Communications. 172 (3): 1230–5. doi:10.1016/0006-291X(90)91580-L. PMID 2244907.
- 1 2 3 4 Kuhn H, Banthiya S, van Leyen K (April 2015). "Mammalian lipoxygenases and their biological relevance". Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids. 1851 (4): 308–30. doi:10.1016/j.bbalip.2014.10.002. PMC 4370320. PMID 25316652.
- 1 2 Yeung J, Holinstat M (July 2011). "12-lipoxygenase: a potential target for novel anti-platelet therapeutics". Cardiovascular & Hematological Agents in Medicinal Chemistry. 9 (3): 154–64. doi:10.2174/187152511797037619. PMC 3171607. PMID 21838667.
- ↑ Nugteren DH (February 1975). "Arachidonate lipoxygenase in blood platelets". Biochimica et Biophysica Acta (BBA) - Lipids and Lipid Metabolism. 380 (2): 299–307. doi:10.1016/0005-2760(75)90016-8. PMID 804329.
- ↑ Krieg P, Fürstenberger G (March 2014). "The role of lipoxygenases in epidermis". Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids. 1841 (3): 390–400. doi:10.1016/j.bbalip.2013.08.005. PMID 23954555.
- ↑ Muñoz-Garcia A, Thomas CP, Keeney DS, Zheng Y, Brash AR (March 2014). "The importance of the lipoxygenase-hepoxilin pathway in the mammalian epidermal barrier". Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids. 1841 (3): 401–8. doi:10.1016/j.bbalip.2013.08.020. PMC 4116325. PMID 24021977.
- 1 2 Pace-Asciak CR (April 2015). "Pathophysiology of the hepoxilins". Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids. 1851 (4): 383–96. doi:10.1016/j.bbalip.2014.09.007. PMID 25240838.
- 1 2 3 4 5 6 Porro B, Songia P, Squellerio I, Tremoli E, Cavalca V (August 2014). "Analysis, physiological and clinical significance of 12-HETE: a neglected platelet-derived 12-lipoxygenase product". Journal of Chromatography B. 964: 26–40. doi:10.1016/j.jchromb.2014.03.015. PMID 24685839.
- 1 2 Weylandt KH (November 2015). "Docosapentaenoic acid derived metabolites and mediators - The new world of lipid mediator medicine in a nutshell". European Journal of Pharmacology. 785: 108–115. doi:10.1016/j.ejphar.2015.11.002. PMID 26546723.
- ↑ Serhan CN (2005). "Lipoxins and aspirin-triggered 15-epi-lipoxins are the first lipid mediators of endogenous anti-inflammation and resolution". Prostaglandins, Leukotrienes, and Essential Fatty Acids. 73 (3–4): 141–62. doi:10.1016/j.plefa.2005.05.002. PMID 16005201.
- ↑ O'Flaherty JT, Hu Y, Wooten RE, Horita DA, Samuel MP, Thomas MJ, Sun H, Edwards IJ (2012). "15-lipoxygenase metabolites of docosahexaenoic acid inhibit prostate cancer cell proliferation and survival". PLOS ONE. 7 (9): e45480. Bibcode:2012PLoSO...745480O. doi:10.1371/journal.pone.0045480. PMC 3447860. PMID 23029040.
- 1 2 Joshi YB, Giannopoulos PF, Praticò D (March 2015). "The 12/15-lipoxygenase as an emerging therapeutic target for Alzheimer's disease". Trends in Pharmacological Sciences. 36 (3): 181–6. doi:10.1016/j.tips.2015.01.005. PMC 4355395. PMID 25708815.
- ↑ "rs2073438". NCBI dbSNP.
- ↑ Quintana LF, Guzmán B, Collado S, Clària J, Poch E (February 2006). "A coding polymorphism in the 12-lipoxygenase gene is associated to essential hypertension and urinary 12(S)-HETE". Kidney International. 69 (3): 526–30. doi:10.1038/sj.ki.5000147. PMID 16514435.
- ↑ Angulo JC, Lopez JI, Dorado JF, Sanchez-Chapado M, Colas B, Ropero S (2016). "A DNA Hypermethylation Profile Independently Predicts Biochemical Recurrence Following Radical Prostatectomy". Urologia Internationalis. 97 (1): 16–25. doi:10.1159/000446446. PMID 27220660. S2CID 12659175.
- ↑ Tang K, Finley RL, Nie D, Honn KV (March 2000). "Identification of 12-lipoxygenase interaction with cellular proteins by yeast two-hybrid screening". Biochemistry. 39 (12): 3185–91. doi:10.1021/bi992664v. PMID 10727209.
- ↑ "rs6502997". NCBI dbSNP.
- ↑ "rs312462". NCBI dbSNP.
- ↑ "rs6502998". NCBI dbSNP.
- ↑ "rs434473". NCBI dbSNP.
- ↑ Witola WH, Liu SR, Montpetit A, Welti R, Hypolite M, Roth M, Zhou Y, Mui E, Cesbron-Delauw MF, Fournie GJ, Cavailles P, Bisanz C, Boyer K, Withers S, Noble AG, Swisher CN, Heydemann PT, Rabiah P, Muench SP, McLeod R (July 2014). "ALOX12 in human toxoplasmosis". Infection and Immunity. 82 (7): 2670–9. doi:10.1128/IAI.01505-13. PMC 4097613. PMID 24686056.
External links
- Human ALOX12 genome location and ALOX12 gene details page in the UCSC Genome Browser.
Further reading
- Yoshimoto T, Arakawa T, Hada T, Yamamoto S, Takahashi E (December 1992). "Structure and chromosomal localization of human arachidonate 12-lipoxygenase gene". The Journal of Biological Chemistry. 267 (34): 24805–9. doi:10.1016/S0021-9258(18)35835-6. PMID 1447217.
- Izumi T, Hoshiko S, Rådmark O, Samuelsson B (October 1990). "Cloning of the cDNA for human 12-lipoxygenase". Proceedings of the National Academy of Sciences of the United States of America. 87 (19): 7477–81. Bibcode:1990PNAS...87.7477I. doi:10.1073/pnas.87.19.7477. PMC 54770. PMID 2217179.
- Funk CD, Furci L, FitzGerald GA (August 1990). "Molecular cloning, primary structure, and expression of the human platelet/erythroleukemia cell 12-lipoxygenase". Proceedings of the National Academy of Sciences of the United States of America. 87 (15): 5638–42. Bibcode:1990PNAS...87.5638F. doi:10.1073/pnas.87.15.5638. PMC 54382. PMID 2377602.
- Flatman S, Morgan A, McDonald-Gibson RG, Davey A, Jonas GE, Slater TF (May 1988). "12-Lipoxygenase activity in human uterine cervix". Prostaglandins, Leukotrienes, and Essential Fatty Acids. 32 (2): 87–94. doi:10.1016/0952-3278(88)90101-9. PMID 3406043.
- Wong PY, Westlund P, Hamberg M, Granström E, Chao PH, Samuelsson B (August 1985). "15-Lipoxygenase in human platelets". The Journal of Biological Chemistry. 260 (16): 9162–5. doi:10.1016/S0021-9258(17)39346-8. PMID 3926763.
- Nakamura M, Ueda N, Kishimoto K, Yoshimoto T, Yamamoto S, Ishimura K (March 1995). "Immunocytochemical localization of platelet-type arachidonate 12-lipoxygenase in mouse blood cells". The Journal of Histochemistry and Cytochemistry. 43 (3): 237–44. doi:10.1177/43.3.7868854. PMID 7868854.
- Hussain H, Shornick LP, Shannon VR, Wilson JD, Funk CD, Pentland AP, Holtzman MJ (January 1994). "Epidermis contains platelet-type 12-lipoxygenase that is overexpressed in germinal layer keratinocytes in psoriasis". The American Journal of Physiology. 266 (1 Pt 1): C243-53. doi:10.1152/ajpcell.1994.266.1.C243. PMID 8304420.
- Arora JK, Lysz TW, Zelenka PS (June 1996). "A role for 12(S)-HETE in the response of human lens epithelial cells to epidermal growth factor and insulin". Investigative Ophthalmology & Visual Science. 37 (7): 1411–8. PMID 8641843.
- Hagmann W, Gao X, Timar J, Chen YQ, Strohmaier AR, Fahrenkopf C, Kagawa D, Lee M, Zacharek A, Honn KV (November 1996). "12-Lipoxygenase in A431 cells: genetic identity, modulation of expression, and intracellular localization". Experimental Cell Research. 228 (2): 197–205. doi:10.1006/excr.1996.0317. PMID 8912711.
- Nakamura M, Yamamoto S, Ishimura K (May 1997). "Subcellular localization of arachidonate 12-lipoxygenase and morphological effect of its overexpression on murine keratinocytes". Cell and Tissue Research. 288 (2): 327–34. doi:10.1007/s004410050818. PMID 9082968. S2CID 23548308.
- Nigam S, Kumar GS, Sutherland M, Schewe T, Ikawa H, Yamasaki Y, Ueda N, Yamamoto S (September 1999). "Metabolic suppression of platelet-type 12-lipoxygenase in human uterine cervix with invasive carcinoma". International Journal of Cancer. 82 (6): 827–31. doi:10.1002/(SICI)1097-0215(19990909)82:6<827::AID-IJC10>3.0.CO;2-Q. PMID 10446449. S2CID 22417409.
- Tornhamre S, Elmqvist A, Lindgren JA (April 2000). "15-Lipoxygenation of leukotriene A4: Studies of 12- and 15-lipoxygenase efficiency to catalyze lipoxin formation". Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids. 1484 (2–3): 298–306. doi:10.1016/S1388-1981(00)00017-2. PMID 10760478.
- Chen BK, Tsai TY, Huang HS, Chen LC, Chang WC, Tsai SB, Chang WC (2003). "Functional role of extracellular signal-regulated kinase activation and c-Jun induction in phorbol ester-induced promoter activation of human 12(S)-lipoxygenase gene". Journal of Biomedical Science. 9 (2): 156–65. doi:10.1159/000048212. PMID 11914583. S2CID 46753449.
- Winer I, Normolle DP, Shureiqi I, Sondak VK, Johnson T, Su L, Brenner DE (October 2002). "Expression of 12-lipoxygenase as a biomarker for melanoma carcinogenesis". Melanoma Research. 12 (5): 429–34. doi:10.1097/00008390-200209000-00003. PMID 12394183. S2CID 27336312.
- Gu J, Wen Y, Mison A, Nadler JL (February 2003). "12-lipoxygenase pathway increases aldosterone production, 3',5'-cyclic adenosine monophosphate response element-binding protein phosphorylation, and p38 mitogen-activated protein kinase activation in H295R human adrenocortical cells". Endocrinology. 144 (2): 534–43. doi:10.1210/en.2002-220580. PMID 12538614.
- Fridman C, Ojopi EP, Gregório SP, Ikenaga EH, Moreno DH, Demetrio FN, Guimarães PE, Vallada HP, Gattaz WF, Dias Neto E (February 2003). "Association of a new polymorphism in ALOX12 gene with bipolar disorder". European Archives of Psychiatry and Clinical Neuroscience. 253 (1): 40–3. doi:10.1007/s00406-003-0404-y. PMID 12664313. S2CID 21064663.