


Gasdermin D (GSDMD) is a protein that in humans is encoded by the GSDMD gene on chromosome 8.[5] It belongs to the gasdermin family which is conserved among vertebrates and comprises six members in humans, GSDMA, GSDMB, GSDMC, GSDMD, GSDME (DFNA5) and DFNB59 (Pejvakin). Members of the gasdermin family are expressed in a variety of cell types including epithelial cells and immune cells. GSDMA, GSDMB, GSDMC, GSDMD and GSDME have been suggested to act as tumour suppressors.[6]
Structure
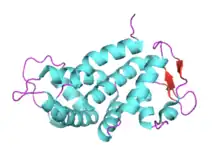
The structure of full-length GSDMD consists of two domains, the 31 kDa N-terminal (GSDMD-N) and 22 kDa C-terminal (GSDMD-C) domains, separated by a linker region. GSDMD-C can be divided into four subdomains and is composed of 10 α-helices and two β-strands, forming a compact globular fold. The linker helix contacts the two helix-repeats which consist of four-helix bundles. The middle domain comprises an antiparallel β-strand and a short α-helix. The first flexible loop of GSDMD-C, which is located between GSDMD-N and the linker helix, stretches out and inserts into the GSDMD-N pocket, stabilizing the conformation of the full-length protein.[7] GSDMD-N forms large transmembrane pores composed of 31 to 34 subunits that allow the release of interleukin-1 (IL-1) family cytokines and drive pyroptosis.[8]
Function
Several current studies have revealed that GSDMD serves as a specific substrate of inflammatory caspases (caspase-1, -4, -5 and -11) and as an effector molecule for the lytic and highly inflammatory form of programmed cell death known as pyroptosis.[9][10] Hence, GSDMD is an essential mediator of host defence against microbial infection and danger signals. The pore-forming activity of the N-terminal cleavage product causes cell swelling and lysis to prevent intracellular pathogens from replicating, and is required for the release of cytoplasmic content such as the inflammatory cytokine interleukin-1β (IL-1β) into the extracellular space to recruit and activate immune cells to the site of infection.[11] GSDMD has an additional potential role as an antimicrobial by binding to cardiolipin (CL) and form pores on bacterial membranes.
Autoinhibition
Under normal conditions, the full-length GSDMD is inactive as the linker loop between the N-terminal and C-terminal domains stabilises the overall conformation of the full-length protein and allows GSDMD-C to fold back on and auto-inhibit GSDMD-N from inducing pyroptosis.[7] Upon interdomain cleavage by inflammatory caspases, the auto-inhibition is relieved and GSDMD-N cytotoxicity is triggered.
Activation
GSDMD can be cleaved and activated by inflammatory caspases through both the canonical and non-canonical pyroptotic pathways.[12]
Canonical inflammasome pathway
Caspase-1, conserved in vertebrates, is involved in the canonical pathway and is activated by canonical inflammasomes such as NLRP3 and NLRC4 inflammasomes, which are multi-protein complexes that are formed upon recognition of specific inflammatory ligands called pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs) in the cytosol by NOD-like receptors (NLRs). Examples include bacterial type 3 secretion system (T3SS) rod protein and flagellin, which are potent activators of NLRC4 inflammasome, and bacterial toxin nigericin that activates NLRP3 inflammasome.[10]
Non-canonical inflammasome pathway
Caspase-11 in mice and its human homolog caspase-4 and -5 are involved in the non-canonical pathway and are activated by directly binding cytosolic lipopolysaccharide (LPS) secreted by gram-negative bacteria.[9]
Upon activation of these caspases, GSDMD undergoes proteolytic cleavage at Asp-275, which is sufficient to drive pyroptosis.[10]
Mechanism
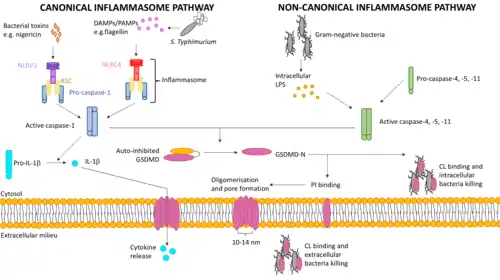
After the proteolytic cleavage, GSDMD-C remains in the cytosol while the N-terminal cleavage product localises to the plasma membrane by anchoring to membrane lipids. GSDMD-N specifically interacts with phosphatidylinositol 4-phosphate [PI(4)P] and phosphatidylinositol 4,5-bisphosphate [PI(4,5)P] on the inner leaflet of mammalian cell membrane strongly, through charge-charge interactions between the negatively-charged head groups of PI and the positively-charged surface on GSDMD-N exposed after cleavage.[13] Hence, collateral damage to tissues during an infection is minimised as the extracellular outer leaflet lacks PI. Lipid binding allows GSDMD-N to insert into the lipid bilayer and induces high-order oligomerisation within the membrane, forming extensive pores with approximately 16 subunits and an inner diameter of 10–14 nm.[7] The osmotic potential is disrupted by pore formation, leading to cell swelling and lysis, the morphologic hallmarks of pyroptosis. The pores also serve as a protein secretion channel to facilitate the secretion of inflammatory cytokines for rapid innate immune response.[14] GSDMD-N can also undergo cytoplasmic distribution and selectively bind to CL on inner and outer leaflets of intracellular bacterial membranes, or be secreted from pyroptotic cells through the pores into the extracellular milieu to target and kill extracellular bacteria.[15]
Clinical significance
Pyroptosis, which can now be defined as gasdermin-mediated necrotic cell death, acts as an immune defence against infection. Hence, failure to express or cleave GSDMD can block pyroptosis and disrupt the secretion of IL-1β, and eventually unable to ablate the replicative niche of intracellular bacteria. Mutation of GSDMD is associated with various genetic diseases and human cancers, including brain, breast, lung, urinary bladder, cervical, skin, oral cavity, pharynx, colon, liver, cecum, stomach, pancreatic, prostate, oesophageal, head and neck, hematologic, thyroid and uterine cancers.[16] Recently, studies have revealed that downregulation of GSDMD promotes gastric cancer proliferation due to the failure to inactivate ERK 1/2, STAT3 and PI3K/AKT pathways, which are involved in cell survival and tumour progression.[17] However, sepsis and lethal septic shock can result from overactivation of pyroptosis.[18]
Gasdermin D also plays a pivotal role in inflammation related MDS development and progression, gasdermin D knockout significantly extends the survival in MDS mouse model.[19] The critical role of GSDMD in pore formation during pyroptosis provides a new avenue for future drug development for treating inflammatory caspase-associated auto-inflammatory conditions, sepsis and septic shock.[16]
Interactions
GSDMD-N has been shown to interact with:[13]
See also
References
- 1 2 3 ENSG00000104518 GRCh38: Ensembl release 89: ENSG00000278718, ENSG00000104518 - Ensembl, May 2017
- 1 2 3 GRCm38: Ensembl release 89: ENSMUSG00000022575 - Ensembl, May 2017
- ↑ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ↑ "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ↑ "Entrez Gene: Gasdermin D". Retrieved 2016-08-26.
- ↑ "GSDMD gasdermin D [Homo sapiens (human)] - Gene - NCBI". www.ncbi.nlm.nih.gov.
- 1 2 3 Kuang S, Zheng J, Yang H, Li S, Duan S, Shen Y, Ji C, Gan J, Xu XW, Li J (October 2017). "Structure insight of GSDMD reveals the basis of GSDMD autoinhibition in cell pyroptosis". Proceedings of the National Academy of Sciences of the United States of America. 114 (40): 10642–10647. Bibcode:2017PNAS..11410642K. doi:10.1073/pnas.1708194114. PMC 5635896. PMID 28928145.
- ↑ Xia S, Zhang Z, Magupalli VG, Pablo JL, Dong Y, Vora SM, et al. (April 2021). "Gasdermin D pore structure reveals preferential release of mature interleukin-1". Nature. 593 (7860): 607–611. Bibcode:2021Natur.593..607X. doi:10.1038/s41586-021-03478-3. PMC 8588876. PMID 33883744. S2CID 233351704.
- 1 2 Dixit V (2015). "Caspase-11 cleaves gasdermin D for non-canonical inflammasome signaling". Nature. 526 (7575): 666–671. Bibcode:2015Natur.526..666K. doi:10.1038/nature15541. PMID 26375259. S2CID 4447176.
- 1 2 3 Shi J, Zhao Y, Wang K, Shi X, Wang Y, Huang H, et al. (October 2015). "Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death". Nature. 526 (7575): 660–5. Bibcode:2015Natur.526..660S. doi:10.1038/nature15514. PMID 26375003. S2CID 4407455.
- ↑ Liu X, Zhang Z, Ruan J, Pan Y, Magupalli VG, Wu H, Lieberman J (July 2016). "Inflammasome-activated gasdermin D causes pyroptosis by forming membrane pores". Nature. 535 (7610): 153–8. Bibcode:2016Natur.535..153L. doi:10.1038/nature18629. PMC 5539988. PMID 27383986.
- ↑ Aglietti RA, Dueber EC (April 2017). "Recent Insights into the Molecular Mechanisms Underlying Pyroptosis and Gasdermin Family Functions". Trends in Immunology. 38 (4): 261–271. doi:10.1016/j.it.2017.01.003. PMID 28196749.
- 1 2 Ding J, Wang K, Liu W, She Y, Sun Q, Shi J, Sun H, Wang DC, Shao F (July 2016). "Pore-forming activity and structural autoinhibition of the gasdermin family". Nature. 535 (7610): 111–6. Bibcode:2016Natur.535..111D. doi:10.1038/nature18590. PMID 27281216. S2CID 4391444.
- ↑ Evavold CL, Ruan J, Tan Y, Xia S, Wu H, Kagan JC (January 2018). "The Pore-Forming Protein Gasdermin D Regulates Interleukin-1 Secretion from Living Macrophages". Immunity. 48 (1): 35–44.e6. doi:10.1016/j.immuni.2017.11.013. PMC 5773350. PMID 29195811.
- ↑ Qiu S, Liu J, Xing F (April 2017). "'Hints' in the killer protein gasdermin D: unveiling the secrets of gasdermins driving cell death". Cell Death and Differentiation. 24 (4): 588–596. doi:10.1038/cdd.2017.24. PMC 5384029. PMID 28362726.
- 1 2 Shi J, Gao W, Shao F (April 2017). "Pyroptosis: Gasdermin-Mediated Programmed Necrotic Cell Death". Trends in Biochemical Sciences. 42 (4): 245–254. doi:10.1016/j.tibs.2016.10.004. PMID 27932073.
- ↑ Wang WJ, Chen D, Jiang MZ, Xu B, Li XW, Chu Y, Zhang YJ, Mao R, Liang J, Fan DM (February 2018). "Downregulation of gasdermin D promotes gastric cancer proliferation by regulating cell cycle-related proteins". Journal of Digestive Diseases. 19 (2): 74–83. doi:10.1111/1751-2980.12576. PMID 29314754.
- ↑ Aziz M, Jacob A, Wang P (November 2014). "Revisiting caspases in sepsis". Cell Death & Disease. 5 (11): e1526. doi:10.1038/cddis.2014.488. PMC 4260746. PMID 25412304.
- ↑ Ren K, Mei Y, Liu Y, Han X, Li E, Bi H, Ji P (2021-11-05). "Gasdermin D Mediates Inflammation-Driven Pathogenesis of the Myelodysplastic Syndromes". Blood. 138 (Supplement 1): 2587. doi:10.1182/blood-2021-149340. ISSN 0006-4971. S2CID 244540484.
Further reading
- Saeki N, Usui T, Aoyagi K, Kim DH, Sato M, Mabuchi T, Yanagihara K, Ogawa K, Sakamoto H, Yoshida T, Sasaki H (March 2009). "Distinctive expression and function of four GSDM family genes (GSDMA-D) in normal and malignant upper gastrointestinal epithelium". Genes, Chromosomes & Cancer. 48 (3): 261–71. doi:10.1002/gcc.20636. PMID 19051310. S2CID 19705164.
This article incorporates text from the United States National Library of Medicine, which is in the public domain.