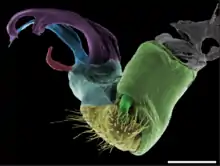
Gonopods are specialized appendages of various arthropods used in reproduction or egg-laying. In males, they facilitate the transfer of sperm from male to female during mating, and thus are a type of intromittent organ. In crustaceans and millipedes, gonopods are modified walking or swimming legs. Gonopods may be highly decorated with elaborate structures which may play roles in sperm competition, and can be used to differentiate and identify closely related species. Gonopods generally occur in one or more pairs, as opposed to the single (un-paired) reproductive organs such as the aedeagus of insects or the penis of harvestmen.
Insects

In insects, gonopods are appendages of the genital segment that may be used in insemination, or that comprise the egg-laying apparatus.[1][2]
Crustaceans
In male decapod crustaceans, gonopods are modified swimming appendages (pleopods). The anterior two pair of pleopods in males are modified for sperm transferring, with differing degree of morphological diversification.[3]
Millipedes

In millipedes, gonopods consist of one or two pairs of often highly modified walking legs in mature males, and are primarily found in members of the subgroup Helminthomorpha—containing most orders and the vast majority of species—where they are located on the seventh body segment consisting of leg pairs 8 and/or 9.[4] Males of the subgroup Pentazonia (which includes the Oniscomorpha (pill millipedes) and Glomeridesmida) lack gonopods but possess enlarged appendages known as telopods at the rear of the body used to firmly hold females during mating.[5] The complex structure of gonopods is a primary method of distinguishing closely related species of millipede,[6] although the terminology used to describe the same structures may vary between authors.[7] The complex morphology of millipede gonopods may be driven by sperm competition or other forms of sexual selection, with some structures serving to scoop out or displace sperm of other males, and others acting to stimulate females into becoming sexually receptive.[5]
Millipede gonopods do not produce sperm directly, but rather gather sperm produced from a gonopore on the base of the third body segment.[8]
Gonopods develop gradually during the growth of an individual. In early developmental stages, all legs are of the walking type, and cannot be used to determine sex. Through successive molts, the walking legs metamorphose into mature gonopods.[8]

| Order | Number of pairs of gonopods/telopods | Leg pair(s) modified![lower-alpha 1] | Notes LP = Leg pair | Image[lower-alpha 2] |
|---|---|---|---|---|
| Callipodida | 1 | 8th | LP9 is reduced | |
| Chordeumatida | 2 | 8th & 9th | Gonopods and other accessory mating legs vary within the four suborders: In Chordeumatidea LP9 sperm-transferring, LP10 is reduced, and LP11 possess coxal glands; in Heterochordeumatidea LP9 sperm-transferring, LP10 reduced, and both LP10 and LP11 with coxal gland; in the Craspedosomatidea and Striariidea, LP8 sperm-transferring, LP9 modified, LP10 and LP11 with coxal glands | |
| Glomerida | 2 (telopods) | Not involved with sperm transfer, only clasping females |  Glomeris troglokabyliana walking legs (A, B) and telopod (C) | |
| Glomeridesmida | 1 (telopods) | |||
| Julida | 2 | 8th & 9th | LP9 sperm-transferring |  Ommatoiulus chambiensis right anterior (1) and posterior (2) |
| Platydesmida | 2 | 9th & 10th | Leg-like; LP 9 sperm-transferring | |
| Polydesmida | 1 | 8th | ||
| Polyxenida | 0 | Indirect fertilization | ||
| Polyzoniida | 2 | 9th & 10th | leg-like; LP 9 sperm-transferring | |
| Siphoniulida | 1 | 8th | ||
| Siphonocryptida | 2 | 9th & 10th | leg-like, LP 9 sperm-transferring | |
| Siphonophorida | 2 | 9th & 10th | leg-like; LP 9 sperm-transferring |  Illacme plenipes, arrow pointing to anterior gonopods |
| Sphaerotheriida | 2 (telopods) |  Sphaeromimus vatovavy posterior telopods | ||
| Spirobolida | 2 | 8th & 9th | LP9 sperm-transferring, LP8 fused into single structure. |  Aphistogoniulus jeekeli anterior (A) and left posterior (B) |
| Spirostreptida | 2 | 8th & 9th | LP8 sperm-transferring | |
| Stemmiulida | 1 | 8th | LP8 sperm-transferring; LP9 reduced |


See also
- Gonopodium, a modified fin for sperm transfer found in some fish
- Claspers, pelvic fins modified for copulation in cartilaginous fish
- Pedipalps, appendages of arachnids involved in sperm-transfer
References
- ↑ Capinera, John L., ed. (2008). Encyclopedia of Entomology (2nd ed.). Dordrecht: Springer. p. 1633. ISBN 9781402062421.
- ↑ Nath, Paras (2007). Illustrated Dictionary of Entomology. Pitam Pura, New Delhi: New India Publishing Agency. p. 183. ISBN 9788189422561.
- ↑ Bauer, Raymond T. (1986). "Phylogenetic Trends in Sperm Transfer and Storage Complexity in Decapod Crustaceans" (PDF). Journal of Crustacean Biology. 6 (3): 313–325. doi:10.2307/1548173. JSTOR 1548173.
- ↑ Sierwald, Petra; Bond, Jason E. (2007). "Current Status of the Myriapod Class Diplopoda (Millipedes): Taxonomic Diversity and Phylogeny". Annual Review of Entomology. 52 (1): 401–420. doi:10.1146/annurev.ento.52.111805.090210. PMID 17163800.
- 1 2 Shear, William A. (1999). "Millipeds". American Scientist. 87 (3): 232–240. Bibcode:1999AmSci..87..232S. doi:10.1511/1999.3.232.
- ↑ Mesibov, Robert. "Gonopods". External Anatomy of Polydesmida. Archived from the original on 9 July 2017. Retrieved 27 October 2013.
- ↑ Car, Catherine; Harvey, Mark (2013). "A review of the Western Australian keeled millipede genus Boreohesperus (Diplopoda, Polydesmida, Paradoxosomatidae)". ZooKeys (290): 1–19. doi:10.3897/zookeys.290.5114. PMC 3677291. PMID 23794855.
- 1 2 Drago, Leandro; Fusco, Giuseppe; Garollo, Elena; Minelli, Alessandro (2011). "Structural aspects of leg-to-gonopod metamorphosis in male helminthomorph millipedes (Diplopoda)". Frontiers in Zoology. 8 (1): 19. doi:10.1186/1742-9994-8-19. PMC 3170261. PMID 21859471.