| Adipose Phospholipase A2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Crystallographic structure of Adipose Phospholipase A2 (AdPLA) | |||||||||
| Identifiers | |||||||||
| EC no. | 3.1.1.4 | ||||||||
| CAS no. | 9001-84-7 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| Gene Ontology | AmiGO / QuickGO | ||||||||
| |||||||||
Group XVI phospholipase A2 also commonly known as adipocyte phospholipase A2 (AdPLA) is an enzyme that in humans is encoded by the PLA2G16 gene.[1][2][3] This enzyme has also been identified as PLA2G16, HRASLS3, HREV107, HREV107-3, MGC118754 or H-REV107-1 from studies on class II tumor suppression but not on its enzymatic properties.[4] AdPLA is encoded by a 1.3 kilobase AdPLA messenger RNA and is an 18 kDa protein. It belongs to a superfamily of phospholipase A2 (PLA2) enzymes and is found primarily in adipose tissue. AdPLA regulates adipocyte lipolysis and release of fatty acids through a G-protein coupled pathway involving prostaglandin and EP3. It has also been reported to play a crucial role in the development of obesity in mouse models.[5]
Enzyme characteristics
AdPLA has been characterized in Group XVI as a separate subgroup of the PLA2 family for its distinct properties from other known PLA2s. It bears similarity to its PLA2 family in phospholipase activity and calcium dependence. Unlike other PLA2 enzymes, AdPLA is expressed predominantly in adipose tissue at higher levels than in the rest of the body, more so in white adipose tissue (WAT) than brown adipose tissue (BAT). Its primary enzymatic function is to catalyze the preferential hydrolysis of phosphatidylcholines at the sn-2 position, generating free fatty acids.
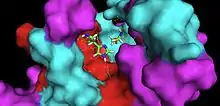
AdPLA contains a membrane-spanning domain on the C-terminus, which localizes intracellularly for phospholipase activity in proximity to cyclooxygenase 1 (COX-1). His-23 and Cys-113 residues have been shown to be essential in AdPLA activity, which differs from the known His/Asp catalytic dyad or Ser/His/Asp catalytic triad of other PLA2 enzymes. Gln-129 and Asn-112 have also been shown to be necessary in catalysis but their role is not known.[2]
AdPLA activity is calcium and pH dependent. Calcium binds to AdPLA and forms a positively charged oxyanion hole to stabilize a negatively charged transition state, similar to other PLA2 active sites. Whereas asparagine binds to calcium in other PLA2 enzymes,[6] the residue that participates in the creation of oxyanion hole in AdPLA has not yet been verified. Optimum AdPLA activity occurs in relatively basic conditions, between pH 7 and 9, to facilitate formation of a histidine-water complex and subsequent fatty acid hydrolysis.[2]
Function


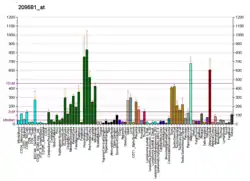
Studies on AdPLA have shown lipolysis regulation following a G-protein coupled pathway in WAT.[5] WAT is responsible for releasing fatty acids from stored triacylglycerol as energy sources for other tissues which is regulated predominately by AdPLA over other phospholipase A2 enzymes. Lipolysis is inversely related to AdPLA activity. AdPLA catalyzes the rate-limiting step, production of arachidonic acid, for the production of prostaglandins, specifically prostaglandin E2 (PGE2). PGE2 enters the signaling pathway binding to G protein-coupled receptor (EP3) which inhibits adenylyl cyclase. Inhibition of adenylyl cyclase decreases the conversion of cyclic AMP (cAMP) from ATP. Lower levels of cAMP decrease the activity of protein kinase A to phosphorylate, thereby activating, hormone-sensitive lipase.[11] The opposite effect can be reached with inactivated AdPLA, decreasing PGE2 concentration and EP3 activity, leading to an increase in cAMP and lipase activity. This mechanism was postulated on the basis that the predominant signaling protein and receptor present in WAT are PGE2 and EP3. These results were based on a mouse model and although they are mammalian cells, it has not been shown to apply to human cells.
Effects on obesity
Obesity has been attributed to adipocyte hypertrophy, where triacylglycerol synthesis exceeds lipolysis, resulting in elevated triacylglycerol storage.[12] Previous studies have associated obesity with endocrine factors and have led pharmacological work toward hormone regulation.[13] Studies on AdPLA deficient mice have shown that the enzyme increased lipolysis in WAT as a result of decreased lipolysis regulation. AdPLA deficiency was shown to reduce adipose tissue mass for mice in both standard and high fat diets. Adipocyte hypotrophy was attributed primarily to reduced triacylglyceride content in WAT from lipolysis, while adipocyte differentiation did not play a role in reduced adipose tissue despite the effects of prostaglandins on adipogenesis.[14] AdPLA deficiency also led to higher oxygen consumption due to the upregulation of genes involved in oxidative metabolism, increasing fatty acid oxidation. One upregulated gene in particular, uncoupling protein-1 (UCP1), has been shown to reduce diet-induced obesity.[15]
Studies on AdPLA deficient and genetically obese mice (leptin deficiency) have also shown similar effects, reduced adipose tissue mass and increased lipolysis by reduction in PGE2 and EP3 activity. Fatty acid oxidation was also found to increase to levels of wild-type mice that were deficient in non-AdPLA deficient obese mice. Body composition also showed a higher percentage of water and lean tissue mass compared to non-AdPLA deficient obese mice.[5]
AdPLA deficiency also demonstrated adverse effects, increasing ectopic triglyceride storage and insulin resistance. Liver enlargement was attributed to higher fatty acid uptake and triacylglycerol content. Insulin stimulated glucose uptake and metabolism were also blunted in AdPLA deficiency, decreasing glycolysis and glycogen synthesis.[5] Despite these side effects, AdPLA is a novel breakthrough in studying autocrine and paracrine action of AdPLA in regulating obesity and fat metabolism. These side effects have triggered new studies to be performed on reduction of AdPLA function as opposed to complete ablation.[16]
References
- ↑ Husmann K, Sers C, Fietze E, Mincheva A, Lichter P, Schäfer R (Oct 1998). "Transcriptional and translational downregulation of H-REV107, a class II tumour suppressor gene located on human chromosome 11q11-12". Oncogene. 17 (10): 1305–12. doi:10.1038/sj.onc.1202060. PMID 9771974.
- 1 2 3 Duncan RE, Sarkadi-Nagy E, Jaworski K, Ahmadian M, Sul HS (Sep 2008). "Identification and functional characterization of adipose-specific phospholipase A2 (AdPLA)". J Biol Chem. 283 (37): 25428–36. doi:10.1074/jbc.M804146200. PMC 2533091. PMID 18614531.
- ↑ "Entrez Gene: HRASLS3 HRAS-like suppressor 3".
- ↑ Sers C, Emmenegger U, Husmann K, Bucher K, Andres AC, Schäfer R (February 1997). "Growth-inhibitory activity and downregulation of the class II tumor-suppressor gene H-rev107 in tumor cell lines and experimental tumors". J. Cell Biol. 136 (4): 935–44. doi:10.1083/jcb.136.4.935. PMC 2132501. PMID 9049257.
- 1 2 3 4 Jaworski K, Ahmadian M, Duncan RE, Sarkadi-Nagy E, Varady KA, Hellerstein MK, Lee HY, Samuel VT, Shulman GI, Kim KH, de Val S, Kang C, Sul HS (February 2009). "AdPLA ablation increases lipolysis and prevents obesity induced by high-fat feeding or leptin deficiency". Nat. Med. 15 (2): 159–68. doi:10.1038/nm.1904. PMC 2863116. PMID 19136964.
- ↑ Six DA, Dennis EA (October 2000). "The expanding superfamily of phospholipase A(2) enzymes: classification and characterization". Biochim. Biophys. Acta. 1488 (1–2): 1–19. doi:10.1016/S1388-1981(00)00105-0. PMID 11080672. S2CID 23717374.
- 1 2 3 GRCh38: Ensembl release 89: ENSG00000176485 - Ensembl, May 2017
- 1 2 3 GRCm38: Ensembl release 89: ENSMUSG00000060675 - Ensembl, May 2017
- ↑ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ↑ "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ↑ Richelsen B (November 1992). "Release and effects of prostaglandins in adipose tissue". Prostaglandins Leukot. Essent. Fatty Acids. 47 (3): 171–82. doi:10.1016/0952-3278(92)90235-B. PMID 1475271.
- ↑ Jaworski K, Sarkadi-Nagy E, Duncan RE, Ahmadian M, Sul HS (July 2007). "Regulation of triglyceride metabolism. IV. Hormonal regulation of lipolysis in adipose tissue". Am. J. Physiol. Gastrointest. Liver Physiol. 293 (1): G1–4. doi:10.1152/ajpgi.00554.2006. PMC 2887286. PMID 17218471.
- ↑ Adan RA, Vanderschuren LJ, la Fleur SE (April 2008). "Anti-obesity drugs and neural circuits of feeding". Trends Pharmacol. Sci. 29 (4): 208–17. doi:10.1016/j.tips.2008.01.008. PMID 18353447.
- ↑ Fajas L, Miard S, Briggs MR, Auwerx J (September 2003). "Selective cyclo-oxygenase-2 inhibitors impair adipocyte differentiation through inhibition of the clonal expansion phase". J. Lipid Res. 44 (9): 1652–9. doi:10.1194/jlr.M300248-JLR200. PMID 12837847.
- ↑ Kopecký J, Hodný Z, Rossmeisl M, Syrový I, Kozak LP (May 1996). "Reduction of dietary obesity in aP2-Ucp transgenic mice: physiology and adipose tissue distribution". Am. J. Physiol. 270 (5 Pt 1): E768–75. doi:10.1152/ajpendo.1996.270.5.E768. PMID 8967464.
- ↑ News Staff. "Disabling AdPLA Enzyme Lets You Eat Anything And Never Get Obese, If..." ION Publications LLC. Retrieved 1 March 2012.
Further reading
- Scanlan MJ, Gordan JD, Williamson B, Stockert E, Bander NH, Jongeneel V, Gure AO, Jäger D, Jäger E, Knuth A, Chen YT, Old LJ (1999). "Antigens recognized by autologous antibody in patients with renal-cell carcinoma". Int. J. Cancer. 83 (4): 456–64. doi:10.1002/(SICI)1097-0215(19991112)83:4<456::AID-IJC4>3.0.CO;2-5. PMID 10508479.
- Siegrist S, Féral C, Chami M, Solhonne B, Mattéi MG, Rajpert-De Meyts E, Guellaën G, Bulle F (2001). "hH-Rev107, a class II tumor suppressor gene, is expressed by post-meiotic testicular germ cells and CIS cells but not by human testicular germ cell tumors". Oncogene. 20 (37): 5155–63. doi:10.1038/sj.onc.1204658. PMC 1865474. PMID 11526504.
- Sers C, Husmann K, Nazarenko I, Reich S, Wiechen K, Zhumabayeva B, Adhikari P, Schröder K, Gontarewicz A, Schäfer R (2002). "The class II tumour suppressor gene H-REV107-1 is a target of interferon-regulatory factor-1 and is involved in IFNgamma-induced cell death in human ovarian carcinoma cells". Oncogene. 21 (18): 2829–39. doi:10.1038/sj.onc.1205377. PMID 11973642.
- Roder K, Kim KH, Sul HS (2002). "Induction of murine H-rev107 gene expression by growth arrest and histone acetylation: involvement of an Sp1/Sp3-binding GC-box". Biochem. Biophys. Res. Commun. 294 (1): 63–70. doi:10.1016/S0006-291X(02)00440-0. PMID 12054741.
- Rual JF, Venkatesan K, Hao T, Hirozane-Kishikawa T, Dricot A, Li N, Berriz GF, Gibbons FD, Dreze M, Ayivi-Guedehoussou N, Klitgord N, Simon C, Boxem M, Milstein S, Rosenberg J, Goldberg DS, Zhang LV, Wong SL, Franklin G, Li S, Albala JS, Lim J, Fraughton C, Llamosas E, Cevik S, Bex C, Lamesch P, Sikorski RS, Vandenhaute J, Zoghbi HY, Smolyar A, Bosak S, Sequerra R, Doucette-Stamm L, Cusick ME, Hill DE, Roth FP, Vidal M (2005). "Towards a proteome-scale map of the human protein-protein interaction network". Nature. 437 (7062): 1173–8. Bibcode:2005Natur.437.1173R. doi:10.1038/nature04209. PMID 16189514. S2CID 4427026.
- Lim J, Hao T, Shaw C, Patel AJ, Szabó G, Rual JF, Fisk CJ, Li N, Smolyar A, Hill DE, Barabási AL, Vidal M, Zoghbi HY (2006). "A protein-protein interaction network for human inherited ataxias and disorders of Purkinje cell degeneration". Cell. 125 (4): 801–14. doi:10.1016/j.cell.2006.03.032. PMID 16713569. S2CID 13709685.
- Nazarenko I, Kristiansen G, Fonfara S, Guenther R, Gieseler C, Kemmner W, Schafer R, Petersen I, Sers C (2006). "H-REV107-1 stimulates growth in non-small cell lung carcinomas via the activation of mitogenic signaling". Am. J. Pathol. 169 (4): 1427–39. doi:10.2353/ajpath.2006.051341. PMC 1698850. PMID 17003497.
- Nazarenko I, Schäfer R, Sers C (2007). "Mechanisms of the HRSL3 tumor suppressor function in ovarian carcinoma cells". J. Cell Sci. 120 (Pt 8): 1393–404. doi:10.1242/jcs.000018. PMID 17374643.