| Hammerschmiede Clay Pit | |
|---|---|
| Stratigraphic range: Serravallian-Tortonian | |
_02.jpg.webp) Image of the clay pit | |
| Type | Fossil site |
| Unit of | Upper Freshwater Molasse |
| Thickness | 25.7 m |
| Lithology | |
| Primary | Silts, sandstone and claystone |
| Location | |
| Coordinates | 47°55′38″N 10°35′28″E / 47.92722°N 10.59111°E |
| Region | |
| Country | |
 Hammerschmiede clay pit (Germany)  Hammerschmiede clay pit (Bavaria) | |
The Hammerschmiede clay pit (German: Tongrube Hammerschmiede) is a fossil bearing locality in Pforzen, Bavaria, Germany most well known for the discovery of Danuvius guggenmosi, the potentially earliest known bipedal ape. With an age of 11.66-11.42 Ma the site dates to the transition between the Middle and Late Miocene epoch, providing an important window into the faunal changes taking place during this time. This correlates to the time just after the Serravallian-Tortonian boundary, and the MN zones 7/8. It is one of the most well-known Miocene sites, with over 15,000 individual fossils and 117 species having been discovered as of 2020. It has been called "the most important German paleontological discovery of the last decades".[1]
History
Hammerschmiede's first fossils were first discovered by the hobby archaeologist Siegulf Guggenmos in 1972.[1] The site became known in academic circles following publications on its rich fauna of micromammals in 1975.[2] Since 2011, the excavations are led by a collaboration between the University of Tübingen and the Senckenberg Museum, and led by Madelaine Böhme, with the help of local volunteers. Excavations were intensified in 2015, following the discovery of a variety of scientifically significant finds.[1] Media attention increased following the description of Danuvius guggenmosi in 2019, its name honoring the discoverer of the site.[3] Nicknamed "Udo", the discovery was described as the oldest known example of an upright gape in Hominidae, with some media even going so far as to call it proof that the cradle of humanity was located in Bavaria.[4] The site is threatened by clay mining.[5]
Geography
The Hammerschmiede clay pit is located within the eastern Allgäu region of Bavaria, within the municipality of Pforzen, at a height above sea level of almost 700 m. It is located just west of the district Hammerschmiede, after which it is named, and lies between the stream Riedgraben to its south, and the river Wertach to its East. Furthermore, the locality Irsee, where fossil plant remains had already been discovered in the 1950s, is located 4.3 km to its southwest.[6] The closest town is Kaufbeuren to its south-east.
Geology and Stratigraphy
The Hammerschmiede locality is part of the Upper Freshwater Molasse within the North Alpine foreland Basin. It is part of the Obere Serie, which includes Germany's youngest Molasse sediments. The 25.7 m thick section is best exposed at the southern end of the clay pit.
The horizontally bedded layers are mostly grey-colored, carbonatic and fine-grained silts, fine sands and claystone. The sedimentary succession can be traced over 150 m in a northern direction, with seven marker beds being traceable across the whole distance. These are three lignite horizons, three sand horizons and one marlstone. The circa 0.45 m thick homogeneous silty-clayey marlstone contains a variety of terrestrial gastropods, and is located in the basal part of the northern profile. Meanwhile, up to 10 mm thick pedogenic carbonate concretions are abundant in the upper 20 cm. A 20 cm thick blackish lignite horizon occurs 1.5 m above this horizon, overlaying 70 cm of yellowish and brownish mottled dark-grey clay. The color of this lignite is platy, it is partly xylitic and can be horizontally replaced by carbonatic peat clay. This succession may be viewed as a complete calcic paleosol, with the marlstone representing the Bk-horizon, the mottled clay the Bt-horizon, and the lignite the A-horizon. Three horizontally continuous bodies of fine-sand are located between the upper and lower lignite horizons, with the lower one being 1.05 m thick, greenish-grey, showing no bedding structures, but containing iron-hydroxide stains. The middle sand horizon, made up of mica rich grey-blue to greyish sand and containing iron-hydroxide stains, is 2.8 m thick at the southern part of the clay pit, and develops into an up to 4 m deep channel at its erosional base in the North. The upper sand horizon, consisting of grey silty fine-sand showing iron-hydroxide stains, is 1.4 m thick and is divided by a 20 cm thick pedogenic bed 0.70 m above its base. This bed, which includes small fossilised roots, consists of mottled clayey and sandy silts. Reworked pedogenic carbonates clay pebbles with a diameter of up to 3 cm occur 30 cm above the base of the horizon.
A 35 cm thick lignite horizon marks the top of the presently exposed sedimentary succession in the Hammerschmiede outcrop. Its basal 25 cm are platy colored and rarely contain xylit, while the upper 10 cm are blackish carbonatic organic clay instead of lignite. It overlays about 10 cm of carbonate-free, grey-greenish and rusty-yellowish mottled sandy and clayey slits. As a 40 cm thick rooted silt horizon, containing abundant powdery, whitish soft carbonate occurs below it, the topmost succession can be interpreted as an immature calcic paleosol.
Directly below this, another, shjallower calcic paleosol is developed. Its A-horizon is formed by 7 cm thick blackish carbonatic peat clay, overlaying the Bt-horizon (5 cm thick grey clay) and Bk-horizon (8 cm thick whitish clay horizon with abundant powdery carbonate). The paleosols rooting system reaches 50 cm below the A-horizon.
Furthermore, several truncated paleosols, which either completely or partly lack the A-horizon, occur especially between the two lower sand horizons. Those are characterized by up to 0.60 m thick grey green-yellowish mottled clayey marls, which contain matrix-supported pedogenic carbonate concretions. Two less developed calcic-paleosols, which are completely lacking A-horizons but clay-rich Btk horizons, make up the basal 4 meters of the succession in the southern profile.[6]
The locality is divided into six stratigraphic layers.
- HAM 1, of an unclear age, but likely correlating with HAM 5.[6]
- HAM 2, of an unclear age.
- HAM 3, is an erosional channel rich in lignitic pebbles near the top of the sequence,[7] of an unclear age, although proposed to be younger than HAM 1.[8]
- HAM 4 is made up of the trough cross stratified sands of the middle sand horizons channel, and has been dated to 11.44 Ma. Well-developed small-scale cross bedding structures and trough cross stratification can be found in the deepest parts, while reworked pedogenic carbonate concretions, pieces of wood and fossils of gastropods and vertebrates are found as channel lag. Its upper part shows well-preserved fossil roots, which can be traced to depths of 2.8 m.[6]
- HAM 5 is the layer providing the most vertebrate fossils, and has been dated to 11.622–11.618 Ma. Its base is located at 681 m above sea level, and made up of 0.3 m grey, sandy marl containing the bivale Margaritifera flabellata and very abundant component-supported pedogenic carbonate concretion, with an average diameter of 10 mm. Above comes a 0.2 m layer of fine-to-medium grained sand, containing both bivalves and caliche pebbles. The following 0.15 m thick section is similar to the basal horizon, although the component-supported pedogenic carbonate concretion are much smaller on average, usually with a diameter of only 1 mm. A trough-shaped fine-to-medium grained sand body, containing well-rounded quartz pebbles with a diameter of up to 35 mm, as well as marble pebbles. Above 0.2 m of green-grey silty clay occurs. This layer contains remains of gastropods and bivalves, flaser bedded finesand laminae, a low number of pedogenic carbonate concretions and very few quartz pebbles. One meter of thick greenish sandy clay to silkish fine sand overlay the 0.85 m thick sand layer. Molluscs are common here, although badly preserved because of partial leaching of their carbonate shells.[6]
- HAM 6, which has been entirely mined down, but based on available information it is likely that it had a lens-like structure, was dominated by proboscidean remains and situated slightly above HAM 4, just below the topmost coal layer. Therefore, its age has been calculated at 11.42 Ma.[9]
Paleoenvironment
The environment of Hammerschmiede was a seasonally dry floodplain of meandering rivers and streams flowing from south to north.[5] HAM 5 represents a rivulet of local origin, with a width of four to five meters, a channel fill thickness of 0.8–1 meter and a mean water depth of ≤ 0.8 m. HAM 4 was a medium-sized river, with a width of roughly 50 m, a channel fill thickness of 4–5 m and a mean water depth of ≤ 4 m. Both channels are asymmetric in cross-section, with a more deeply incised outer bank and a shallower slip-off slope. Based on grain sizes, the flow velocity of both channels has been estimated as low to very low.[10] However, the presence of an autochthonous freshwater mollusc fauna made up out of Pisidium, Margaritifera, Sphaerium and Bithynia suggests well-oxygenated waters, with moderate flow.[11] The presence of considerable amounts of water are furthermore supported by the abundance of fish and turtle fossils, as well as swamp plants.[12] Pebbles of alpine origin are not present, but reworked marine plankton from the Upper Marine Molasse is. Therefore, the catchment area of these rivers did not include alpine areas. Instead, they likely had their source in the Faltenmolasse just a few kilometers south of Hammerschmiede.[5]
The paleoclimate of the locality was subtropical and considerably warmer and drier than that of the region today,[5] with annual average temperatures of perhaps over 20 °C.[10] Estimates based on the herpetofauna suggest mean annual precipitation (MAP) of 974 ± 256 mm for HAM 1, 1008 ± 257 mm for HAM 2 and 1196 ± 263 mm for HAM 3,[13] while a MAP of 858 ± 147 mm has been estimated for the calcic paleosol with an estimated age of 11.64 ma.[6] Another study estimates mean warm month temperatures of 24.7-27.9 °C, warm month precipitation of 79–172 mm and an average Köppen Aridity Index of 25.4.[14]
All this suggests a seasonally dry climate, with swamps developing in the riparian regions during wetter periods. The high temperatures and comparatively low precipitations suggest a significant seasonal water stress, which is furthermore supported by the abundant pedogenic carbonate concretions, indications of forest fires and an as of yet unpublished stable isotope analysis, which showcases the enriched δ13C values of a C3 vegetation under very strong water stress.[5][6] The large number of scansorial species suggests that the site was forested.[15]
Biostratigraphy and Paleoecology
Hammerschmiede dates to the latest stages of MN7/8, overlapping with the Astaracian-Vallesian boundary. Notable is the presence of Kretzoiarctos beatrix and Sinohippus sp., which link it with the slightly older locality Nombrevilla 2. Another taxon connecting Hammerschmiede to this Spanish locality is Galerix cf. exilis, thought to have gone extinct in Central Europe during earlier in the Middle Miocene, which likely reemmigrated from the Iberian peninsula to Germany during this time.[7] Species such as these suggest that Hammerschmiede, and contemporary sites, may represent the last European faunas with Middle Miocene character. In slightly younger deposits, for example those correlating to the Mytilopsis ornitopsis zone (11.5 to 11.4 Ma, Vienna Basin) many lineages of large middle Miocene mammals have already been replaced with new immigrants,[6] such as Hipparionine horses which define the MN9 zone.[8] However, more typical Late Miocene taxa such as the large bovid Miotragocerus and the cricetid Microtocricetus molassicus are also present.[6] Indeed, the occurrence of this cricetid originally led researchers to the conclusion that Hammerschmiede should be dated to MN9.[8] Furthermore, Hammerschmiede shows both the first and last occurrences of a number of taxa, such as Eomellivora and Laphyctis, respectively.[16]
The carnivoran assemblage of Hammerschmiede is the third richest known from Miocene Europe, with 28 different species. 13 sympatric small carnivorans are known from HAM 5 alone, a diversity comparable with the African tropics.[16] The comparison with six other fossil assemblages (Sansan, Steinheim, La Grive-Saint Alban, Rudabánya, Can Ponsic and Can Llobateres 1) shows some marked differences between the carnivoran guild of those sites and Hammerschmiede. Amphicyonids, extremely common at the other localities, and barbourofelids, which are a regular presence, are only known from infrequent and fragmentary material at Hammerschmiede. Herpestids and lophocyonids, which are known from Sansan and La Grive, are completely absent, while Potamotherium and phocids are not known from the other localities.[15] Mustelids are the most diverse family, with 11 species, making up 39% of all Carnivora present at the locality. Viverrids are only presented by 3 species (11%), but make up 21% of all individuals found. However, there is a marked difference between their abundance in HAM 4 (36%) and HAM 5 (4%). HAM 4 generally shows a lower diversity of large, scansorial and hypercarnivorous carnivorans in comparison with HAM 5, although the number of semiaquatic species is much higher with four. Notably, it is the only locality in which Lartetictis and Paralutra are known to have coexisted, despite the suggestion that the latter genus gradually replaced the former.[16] Other likely competitors include Viverrictis-Circamustela and Semigenetta sansaniensis-"Martes" sansaniensis. Larger species with overlapping ecological roles include the metailurine and barbourofelid. The latters shows more derived sabertooth features, suggesting it likely preyed on larger animals than the felid, which probably attacked medium-sized prey from trees. The coexistence of all these competing taxa can be explained by the existence of significant resources.[15][16] Niche partitioning is also seen among the proboscideans, with Tetralophodon being a mixed feeder, and Deinotherium a browser. As Gomphotherium is absent from this locality, it has been suggested that it correlates to the transition period from Middle Miocene faunas dominated by trilophodont gompothere taxa to the Late Miocene ones dominated by tetralophodont species.[17]
Paleofauna
Color key
|
Notes Uncertain or tentative taxa are in small text; |
Arthropoda
| Name | Species | Layer | Material | Image |
|---|---|---|---|---|
| Ephemeroptera indet. | Larvae |
Molluscs
Bivalvia
| Name | Species | Layer | Material | Notes | Image |
|---|---|---|---|---|---|
| Margaritifera/Pseudunio[11] | M. flabellata | HAM 4[6]
HAM 5 |
Unionid freshwater mussel, the most abundant bivalve of the Upper Freshwater Molasse, forming dense, monospecific communities in clean sand or gravel bottoms. | ||
| Sphaerium[11] | S. rivicola | HAM 4[6] | fragmentary valves | First record from the Upper Freshwater Molasse. Inhabits muddy sand near riverbanks, with moderately strong currents, well-oxygenated water, but little submerged vegetation. |  Sphaerium rivicola |
| Pisidium[11] | P. amnicum | HAM 4[6] | Two contiguous and a great number of individual valves | Inhabits sand in well-oxygenated waters, preferring strong currents and hard water |  Pisidium amnicum |
Gastropoda
| Name | Species | Layer | Material | Notes | Image |
|---|---|---|---|---|---|
| Bithynia[11] | C. sp. 1 | HAM 4[6] | Four shells of sub-adult individuals and a
single operculum. |
Generalist inhabiting limnic and fluval waters. | |
| B. sp. 2 | HAM 4[6] | Over 70 opercula | |||
| Borysthenia[11] | B. sp. | HAM 4[6] | two almost complete shells | Earliest record of the genus. Inhabits sandy and muddy substrate near river banks. | |
| Triptychia[6] | Terrestrial snail belonging to the family Filholiidae | ||||
| Helicoidea indet.[6] | Terrestrial snail, possibly belonging to the genus Pseudochloritis |
Fish
| Name | Species | Layer | Material | Notes | Image |
|---|---|---|---|---|---|
| Esox[3] | E. sp. | HAM 1
HAM 3 HAM 5[18] |
 Northern Pike (Esox lucius) | ||
| Silurus[3] | S. joergi[19] | HAM 1
HAM 3 |
Medium-sized catfish, the oldest member of the genus in Europe |  Extant Silurus lithophilus | |
| Cobitis[6] | C. sp. | HAM 1
HAM 3 HAM 5[18] |
|||
| Tinca[3] | T. sp. | HAM 1
HAM 3 HAM 5[18] |
 Extant tench (Tinca tinca) | ||
| Palaeoleuciscus[3] | P. sp. | HAM 1
HAM 3 HAM 5[18] |
 Paleoleuciscus oeningensis | ||
| Leuciscus[3] | L. sp. | HAM 1
HAM 3 HAM 5[18] |
 Common dace (Leuciscus leuciscus) | ||
| Barbus[3] | B. sp. | HAM 5 |  Barbus plebejus | ||
| Gobius[3] | G. sp. | HAM 1
HAM 3 HAM 5[18] |
_1.jpg.webp) Gobius niger | ||
| Perca[3] | P. sp. | HAM 1
HAM 3 HAM 5[18] |
 European perch (Perca fluviatilis) | ||
| Sciaenidae indet.[3] | HAM 5 |
Amphibia
Anura
| Name | Species | Layer | Material | Notes | Image |
|---|---|---|---|---|---|
| Eopelobates[3] | E. sp. | HAM 3[21]
HAM 5 |
 Eopelobates | ||
| Palaeobatrachus[6] | P. sp. | HAM 3[21]
HAM 5 |
 Life reconstruction of Palaeobatrachus gigas | ||
| Latonia[3] | L. gigantea | HAM 1[18]
HAM 3[21] HAM 5 |
|||
| Pelophylax[3] | P. sp. | HAM 1[18]
HAM 3[21] HAM 5 |
|||
| Hyla[6] | H. sp. | HAM 1[18]
HAM 5 |
|||
| Bufotes[3] | B. cf. viridis | HAM 3[18]
HAM 5 |
Referred to as Bufo cf. viridis |  Bufotes viridis |
Urodela
| Name | Species | Layer | Material | Notes | Image |
|---|---|---|---|---|---|
| Batrachosauroididae indet.[3] | HAM 1[18]
HAM 3[21] HAM 5 |
Occasionally spelled "Batrachosauroidae" | |||
| aff. Palaeoproteus[21] | aff. P. sp. | HAM 1
HAM 3 |
Batrachosauroidid salamander | ||
| Scapherpetontidae indet.[3] | HAM 3[18]
HAM 5 |
||||
| Andrias[3] | A. scheuchzeri | HAM 4[6]
HAM 5 |
Giant salamander |  Andrias scheuchzeri | |
| Mioproteus[3] | M. aff. weizei | HAM 1[18]
HAM 3[21] HAM 5 |
Fossil salamander related to the olm |  Life reconstruction of Mioproteus wezei | |
| Chelotriton[3] | C. paradoxus | HAM 1[18]
HAM 3[21] HAM 5 |
A newt |  Chelotriton paradoxus | |
| Triturus[6] | T. roehrsi | HAM 1[18]
HAM 3[21] HAM 5 |
A newt | ||
| T. aff. montadoni | HAM 3[21]
HAM 5 |
Reptiles
Choristodera
| Name | Species | Layer | Material | Notes | Image |
|---|---|---|---|---|---|
| Lazarussuchus[3] | L. sp. | HAM 2[18]
HAM 3[21] HAM 5 |
The youngest fossils referred to this taxon | 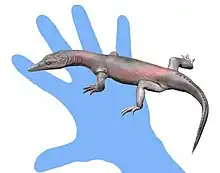 Life reconstruction of Lazarussuchus inexpectatus |
Testudines
| Name | Species | Layer | Material | Notes | Image |
|---|---|---|---|---|---|
| Trionyx[1] | T. sp. | HAM 1[21]
HAM 5 |
Plate fragments[22] | A softshell turtle | |
| Chelydropsis[3] | C. sp. | HAM 1[21]
HAM 4[6] HAM 5 |
Over a thousand remains, including eggs and a well-preserved skull. | Most commonly found tetrapod at the locality. With a length of 70 cm the largest European turtle of its time. Likely molluscivorous chelydrid[23] | |
| Titanochelon[3] | T. sp. | HAM 5 | A large tortoise | ||
| Testudo[3] | T. sp. | HAM 1[21]
HAM 5 |
A tortoise | ||
| Clemmydopsis[3] | C. sp. | HAM 1[21]
HAM 4[6] HAM 5 |
Carapace of a juvenile | A swamp turtle | |
| Mauremys | M. sarmatica | HAM 5 | A swamp turtle |
Squamata
| Name | Species | Layer | Material | Notes | Image |
|---|---|---|---|---|---|
| Lacerta[6][13] | L. sp. 1 | HAM 1
HAM 2 HAM 3 HAM 5 |
|||
| L. sp. 2 | HAM 1
HAM 5 |
||||
| Pseudopus[3][13] | P. pannonicus | HAM 1
HAM 2 HAM 3 HAM 5 |
A great number of scales. | Anguid, with a length of up to two meters the largest lizard present[24] | .jpg.webp) Sheltopusik (Pseudopus apodus) |
| Ophisaurus[3][13] | O. sp. | HAM 1
HAM 2 HAM 3 HAM 5 |
Anguid lizard | ||
| Chalcides[6] | C. sp. | HAM 1[13] HAM 5 | A skink | ||
| Scincidea indet.[13] | HAM 2
HAM 3 |
||||
| Amphisbaenidae indet.[3] | HAM 1[21]
HAM 5 |
||||
| Colubrinae sp. 1[3] | HAM 1[18]
HAM 5 |
||||
| Colubrinae sp. 2[3] | HAM 1[18]
HAM 5 |
||||
| Natricinae sp. 1[3] | HAM 1[18]
HAM 5 |
||||
| Natricinae sp. 2[3] | HAM 5 | ||||
| Erycinae indet. | HAM 5 |
Aves
| Name | Species | Layer | Material | Notes | Image |
|---|---|---|---|---|---|
| Accipitriformes indet.[3] | HAM 5 | One or two species[10] | |||
| Alcedinidae indet.[10] | About the size of the Collared Kingfisher. First fossil record of the family in Europe. | ||||
| Allgoviachen[25] | A. tortonica | HAM 4
HAM 5 |
Nearly complete leg | Anatid comparable in size to an Egyptian goose | |
| cf. Mioquerquedula[25] | cf. M. sp. | HAM 4 | Three wing and pectoral girdle bones | Tiny anatid even smaller than the smallest extant species | |
| Anatidae sp. 1[25] | HAM 4 | Undetermined medium-sized anatid | |||
| Anatidae sp. 2[25] | HAM 4 | Undetermined medium-sized anatid | |||
| Anhinga[10] | A. pannonica | HAM 4
HAM 5 |
Various postcranial remains | Large sized darter | 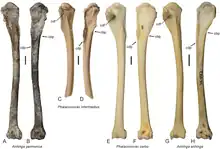 Humerus of Anhinga pannonica |
| Galliformes indet.[3] | HAM 5 | ||||
| Galliformes indet. 2[10] | |||||
| Gruidae indet.[26] | HAM 4 | Dorsal portions of beak and neurocranium | Very large crane, with a skull similar in size and shape to the Siberian crane | ||
| Passeriformes indet.[10] | About the size of the Eurasian Magpie | ||||
| Phalacrocoracidae indet.[10] | HAM 4 | Small cormorant |
Mammalia
Artiodactyla
| Name | Species | Layer | Material | Notes | Image |
|---|---|---|---|---|---|
| Listriodon[3] | L. splendens | HAM 5 | A listriodontine suid |  Listriodon splendens | |
| Parachleuastochoerus[3] | P. steinheimensis | HAM 5 | Perfectly preserved skull[27] | A tetraconodontine suid | |
| Dorcatherium[3] | D. naui | HAM 5 | Large tragulid with pronounced sexual dimorphism[28] |  Dorcatherium | |
| Miotragocerus[3] | M. monacensis | HAM 4[29]
HAM 5 |
Partial skull, postcranial material, teeth, horn cores[30] | Large, boselaphine bovid. Youngest known fossils from this species. |  Miotragocerus pannoniae |
| Boselaphinae indet.[3] | HAM 5 | ||||
| Euprox[3] | E. furcatus | HAM 5 | A cervid | ||
| Muntiacini indet.[3] | HAM 5 | ||||
| Moschidae indet.[3] | HAM 5 |
Carnivora
Caniforma
| Name | Species | Layer | Material | Notes | Image |
|---|---|---|---|---|---|
| Alopecocyon[15] | A. goeriachensis | HAM 5 | teeth, mandible | Small, scansorial simocyonine ailurid[16] | |
| Simocyoninae indet.[15] | HAM 4 | molar | Rather large possibly belongs to Protursus[16] | ||
| Amphicyonidae indet.[15] | HAM 5 | isolated teeth | Considerably larger than Amphicyon major | ||
| Kretzoiarctos[15] | K. beatrix | HAM 4
HAM 5 |
teeth | Ailuropodine ursid, opportunistic feeder weighing around 100 kg | 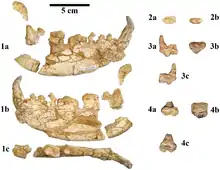 Fossil lower jaws of Kretzoiarctos beatrix |
| Phocidae indet.[15] | HAM 4
HAM 5 |
teeth | Piscivore, slightly larger than the harbor seal | ||
| Proputorius[15] | P. pusillus | HAM 1 | teeth | Tiny (<1 kg), hypocarnivorous mephitid[16] | |
| P. sansaniensis | HAM 1
HAM 5 |
teeth | Small (1–3 kg), hypocarnivorous mephitid[16] | ||
| Palaeomeles[15] | P. pachecoi | HAM 5 | mandible, teeth | Badger-like mephitid, slightly smaller than Meles[16] | |
| Circamustela[15] | C. hartmanni | HAM 1
HAM 4 HAM 5 |
mandible, teeth | Small, hypercarnivorous gulonine[16] | |
|
M. munki |
HAM 4 HAM 5 |
right hemimandible |
smaller than "M". sansaniensis[16] | ||
|
M. sansaniensis |
HAM 4
HAM 5 |
skull | Rather large (3 kg) marten[16] | ||
|
M. sp. |
HAM 1 |
teeth | Small, <1 kg[16] | ||
| Laphyctis[15] | L. mustelinus | HAM 4 | molar | Almost wolverine sized gulonine, potentially belongs to the Genus Ischyrictis[16] | |
| Gulolinae indet.[15] | HAM 5 | left hemimandible and left femur | Hypocarnivore, weighing 5 kg[16] | ||
| Eomellivora[15] | E. moralesi | HAM 5 | right hemimandible, teeth | Oldest occurrence of the genus, slightly larger than a wolverine[16] | |
| Lartetictis[15] | L. cf. dubia | HAM 4 | left hemimandible, teeth | Semiaquatic mustelid[16] | |
| Paralutra[15] | P. jaegeri | HAM 4
HAM 5 |
teeth | Small (<10 kg) otter[16] | |
| Vishnuonyx[15] | V. neptuni | HAM 4 | mandible, teeth | Rather large, piscivorous (10–15 kg) otter[31] | |
| Trocharion[15] | T. albanense | HAM 4
HAM 5 |
skull, teeth | House cat sized leptarcine, hypocarnivore[16] | |
| Potamotherium[15] | P. sp. | HAM 4
HAM 5 |
isolated teeth | Semiaquatic, piscivorous carnivoran of unclear taxonomic status, comparable in size to the smooth-coated otter[16] |  Life restoration of Potamotherium miocenicum. |
Feliforma
| Name | Species | Layer | Material | Notes | Image |
|---|---|---|---|---|---|
| Barbourofelidae indet.[15] | HAM 5 | right distal humerus | A very large form (>100 kg). Similar fossils have been found in Spain. | ||
| Metailurini indet.[15] | HAM 5 | right upper canine | Potentially a connection between Pseudaelurus and Metailurus. Large, similar in size to M. major. | ||
| Pseudaelurus[15] | P. quadridentatus | HAM 5 | teeth | Leopard-sized felid | 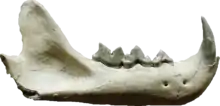 Mandible of Pseudaelurus |
|
L. vallesiensis |
HAM 5 |
Not mentioned in the review of Hammerschmiede carnivorans[15] |
|||
| Hyaenidae indet.[15] | incisor | Very large form (>100 kg). Potentially either belongs to Dinocrocuta or an early crocutoid[9] | |||
| Thalassictis[15] | T. montadai | HAM 4?
HAM 5 HAM 6 |
teeth | Wolf-like ictitheriine hyena[9] | |
| Semigenetta[15] | S. grandis | HAM 4 | teeth | Large (ca. 10 kg) viverrid, hypercarnivore with a similar dietary niche as jackals[16][32] | |
| S. sansaniensis | HAM 1
HAM 4 HAM 5 |
various remains | Most common carnivoran present. Scansorial omnivore leaning towards meat, with a dietary niche comparable to the small Indian civet. Weight of 1.5–4 kg, individuals from Hammerschmiede closer towards the lower end.[16] | ||
| Viverrictis[15] | V. modica | HAM 4
HAM 5 |
teeth | Very small (<1 kg), scansorial, insectivorous viverrid[16] |
Chiroptera
| Name | Species | Layer | Material | Notes | Image |
|---|---|---|---|---|---|
| Chiroptera indet.[3] | various species | HAM 5 |
Eulipotyphla
| Name | Species | Layer | Material | Notes | Image |
|---|---|---|---|---|---|
| Plesiosorex[3] | P. schaffneri | HAM 1[2]
HAM 5 |
A plesiosoricid | ||
| Crusafontina[3] | C. exulta | HAM 1
HAM 3[33] HAM 5 |
Maxilla and teeth | A member of the obscure, highly specialized shrew tribe Anourosoricini[34] | |
| Dinosorex[3] | D. sp. nov. | HAM 5 | A soricid | ||
| Paenelimnoecus[3] | P. crouzeli | HAM 5 | A soricid | ||
| Soricidae indet. 1[3] | HAM 5 | ||||
| Soricidae indet. 2[3] | HAM 5 | ||||
| Desmanella[3] | D. sp. | HAM 5 | A talpid related to shrew moles | ||
| Gehardstorchia[3] | G. quinquecuspidata | HAM 1[2]
HAM 5 |
A talpid | ||
| Proscapanus[3] | P. sp. | HAM 5 | A talpid | ||
| Talpa[3] | T. sp. | HAM 5 | A talpid |  Talpa europaea | |
| Erinaceinae indet.[3] | HAM 5 | ||||
| Galerix[7] | G. cf. exilis | HAM 3 | molars | A galercine, last known Central European occurrence | |
| Plesiodimylus[3] | P. johanni | HAM 5 | A dimylid | ||
| Metacordylodon[3] | M. schlosseri | HAM 5 | A dimylid |
Lagomorpha
| Name | Species | Layer | Material | Notes | Image |
|---|---|---|---|---|---|
| Eurolagus[3] | E. fontannesi | HAM 1[2]
HAM 5 |
An ochotonid | ||
| Prolagus[3] | P. oeningensis | HAM 5 | An ochotonid |  Prolagus oeningensis |
Primates
| Name | Species | Layer | Material | Notes | Image |
|---|---|---|---|---|---|
| Danuvius[3] | D. guggenmosi | HAM 5 | Partial skeleton, and a variety of additional remains | A small hominoid, potentially capable of bipedalism, although that has been disputed[35] | |
| Pliopithecidae indet.[3] | HAM 5 | Teeth[5] | New species |
Perissodactyla
| Name | Species | Layer | Material | Notes | Image |
|---|---|---|---|---|---|
| Ancylotherium[3] | A. sp. | HAM 5 | A schizotheriine | ||
| Aceratherium[2] | A. sp. | HAM 1
HAM 4[6] |
A hornless rhinoceros | ||
| Hoploaceratherium[3] | H. belvederense | HAM 5 | A one-ton heavy, folivorous rhinoceros with a small horn[36] | ||
| Sinohippus[3] | S. sp. | HAM 5 | A folivorous, anchitheriine equid |
Proboscidea
| Name | Species | Layer | Material | Notes | Image |
|---|---|---|---|---|---|
| Deinotherium[3] | D. levius[17] | HAM 4
HAM 5 |
Remains of several juveniles and one adult | A deinotheriine proboscidean | -Deinotherium_Giganteum.jpg.webp) Life restoration of Deinotherium giganteum, Naturhistorisches Museum Mainz |
| Tetralophodon[3] | T.. longirostris | HAM 5
HAM 6[5] |
Partial skeleton of an adult, remains of a juvenile[37] | An elephantoid proboscidean |  Life restoration of Tetralophodon |
Rodentia
| Name | Species | Layer | Material | Notes | Image |
|---|---|---|---|---|---|
| Anomalomys[3] | A. gaudryi | HAM 1[2]
HAM 5 |
An anomalomyid rodent | ||
| Albanesia[3] | A. grimmi | HAM 5 | A flying squirrel | ||
| Blackia[3] | B. miocaenica | HAM 5 | A flying squirrel | ||
| Miopetaurista[2] | M. crusafonti | HAM 1 | A flying squirrel, synonymous with M. albanensis | ||
| Petauristinae indet.[3] | HAM 5 | ||||
| Spermophilinus[3] | S. bredai | HAM 1[2]
HAM 3[38] HAM 5 |
A xerine squirrel | ||
| Chalicomys[3] | C. jaegeri | HAM 4[6]
HAM 5 |
A semiaquatic castorid[39] | ||
| Steneofiber[40] | S. depereti | HAM 4
HAM 5 |
Teeth | A medium sized, semiaquatic castorid | |
| Trogontherium[3] | T. minutus | HAM 1[2]
HAM 4[6] HAM 5 |
A semiaquatic castorid, with similarities to Ondatra[41] Also known as Euroxenomys | ||
| Eliomys[3] | E. sp. | HAM 5 | A glirid rodent |  Eliomys quercinus | |
| Glirulus[3] | G. conjunctus | HAM 5 | A glirid rodent |  Glirulus japonicus | |
| Microdyromys[3] | M. complicatus | HAM 5 | A glirid rodent, synonymous with M. miocaenicus[42] | ||
| Muscardinus[3] | M. hispanicus | HAM 5 | A glirid rodent | ||
| Myoglis[3] | M. meini | HAM 1[2]
HAM 5 |
A glirid rodent | ||
| Eomyops[3] | E. catalaunicus | HAM 1
HAM 2 HAM 3[43] HAM 5 |
great number of isolated teeth | An eomyid rodent | |
| Keramidomys[3] | K. sp. | HAM 5 | An eomyid rodent | ||
| Collimys[3] | C. hiri | HAM 1
HAM 3[8] HAM 5 |
Great number of isolated teeth | A cricetid rodent | |
| C. dobosi[8] | Isolated teeth | A cricetid rodent | |||
| Democricetodon[3] | D. nov. sp. | HAM 1[2]
HAM 5 |
A cricetid rodent | ||
| Eumyarion[3] | E. latior | HAM 5 | A cricetid rodent | ||
| Megacricetodon[3] | M. minutus | HAM 1[2]
HAM 5 |
A cricetid rodent | ||
| Microtocricetus[3] | M. molassicus | HAM 5 | A cricetid rodent |
Paleoflora
| Name | Species | Notes | Image |
|---|---|---|---|
| Alnus[44] | A. sp. |  | |
| Asteraceae[44] | |||
| Betula[44] | B. sp. |  | |
| Buxus[44] | B. sp. |  | |
| Caldesia[12] |  | ||
| Carex[12] |  | ||
| Carpinus[44] | A. sp. | .JPG.webp) | |
| Carya[44] | C. sp. |  | |
| Celtis[44] | C. sp. |  | |
| Ceratophyllum[12] |  | ||
| Decodon[12] |  | ||
| Ephedripites[44] | E. sp. | ||
| Ericaceae[44] | |||
| Extrapunctatosporis[44] | E. miocaenicus | ||
| Fagus[44] | F. sp. | _in_Humlamaden_1516.jpg.webp) | |
| Foveotriletes[44] | F. sp. | ||
| Hydrosporis[44] | H. levis | ||
| Ilex[44] | I. sp. | .jpg.webp) | |
| Inaperturopollenites[44] | I. concedipites | ||
| Juglans[44] | J. sp. |  | |
| Laevigatosporites[44] | L. sp. | ||
| Leiotriletes[44] | L. sp.
L. neddenioides L. wolffi brevis L. wolffi wolffi |
||
| Liquidambar[44] | L. sp. |  | |
| Lonicera[44] | L. sp. |  | |
| Magnoliaepollenites[44] | M. sp. | ||
| Mneme | |||
| Momipites[44] | M. punctatus | ||
| Monogemmites[44] | M. pseudosetarius | ||
| Monoleiotriletes[44] | M. gracilis | ||
| Myrica[44] | M. sp. | female.JPG.webp) | |
| Nymphaea[12] |  | ||
| Nyssa[44] |  | ||
| Oleaceae[44] | |||
| Onagraceae[44] | |||
| Perinomonoletes[44] | P. sp. | ||
| Piceapollis[44] | P. sp. | ||
| Pityosporites[44] | P. alatus
P. insignis P. labdacus P. pristinipollinius |
||
| Platycarya[44] | P. sp. |  | |
| Poaceae[44] | |||
| Podocarpidites[44] | P. sp. | ||
| Polycolpopollenites[44] | P. hexaradiatus | ||
| Potamogeton[12] |  | ||
| Proserpinaca[12] |  | ||
| Pterocarya[44] |  | ||
| Quercoidites[44] | Q. henrici
Q. microhenrici |
||
| Quercus[44] | Q. sp. |  | |
| Retitriletes[44] | R. sp. | ||
| Salix[44] | S. sp. |  | |
| Sambucus[12] |  | ||
| Sapotaceae[44] | |||
| Sciadopitys[44] | S. sp. |  | |
| Sequoiapollenites[44] | S. sp. | ||
| Sparganiaceaepollenites[44] | S. polygonalis | ||
| Sparganium[12] |  | ||
| Spirematospermum[12] | Belongs to the order Zingiberales and inhabited water-associated habitats[45] | ||
| Sphagnum[44] | S. sp. |  | |
| Stratiotes[12] |  | ||
|
Swida[12] |
Junior synonym of Cornus |
 | |
| Tiliaceae[44] | |||
| Tricolpopollenites[44] | T. sp.
T. liblarensis fallax T. cingulum T. cingulum pusillus T. megaexactus bruehlensis T. megaexactus exactus T. pseudocingulum T. pulcher T. wackersdorfensis T. multivallatus |
||
| Tsuga[44] | T. sp. |  | |
| Ulmus[44] | U. sp. |  | |
| Verrucatosporites[44] | V. sp. | ||
| Vitis[12] |  | ||
| Zelkova[44] | Z. sp. |  |
References
- 1 2 3 4 Lechner, Thomas; Böhme, Madelaine. "Eine Fundgrube für Paläontologen - Die Grabungen in der Hammerschmiede bei Pforzen, Lkr. Ostallgäu". Bayerische Archäologie.
- 1 2 3 4 5 6 7 8 9 10 11 12 Helmut Mayr, Volker Fahlbusch: Eine unterpliozäne Kleinsäugerfauna aus der Oberen Süßwasser-Molasse Bayerns. In: Mitteilungen der Bayerischen Staatssammlung für Paläontologie und Historische Geologie. Band 15, 1975, S. 91–111, Volltext (PDF) Archived 2021-06-29 at the Wayback Machine.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 Madelaine Böhme Nikolai Spassov, Jochen Fuss, Adrian Tröscher, Andrew S. Deane, Jérôme Prieto, Uwe Kirscher, Thomas Lechner, David R. Begun: A new Miocene ape and locomotion in the ancestor of great apes and humans. In: Nature. Band 575, 2019, S. 489–493, doi:10.1038/s41586-019-1731-0.
- ↑ Sessler, Stefan (2019). "Die Wiege der Menschheit liegt in Pforzen". Merkur.de. Retrieved 1 November 2022.
- 1 2 3 4 5 6 7 "Hammerschmiede - Das Ost-Allgäu vor 11.5 Millionen Jahren". uni-tuebingen.de. Archived from the original on 1 November 2022.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 Kirscher, U.; Prieto, J.; Bachtadse, V.; Aziz, H. Abdul; Doppler, G.; Hagmaier, M.; Böhme, M. (2016-08-01). "A biochronologic tie-point for the base of the Tortonian stage in European terrestrial settings: Magnetostratigraphy of the topmost Upper Freshwater Molasse sediments of the North Alpine Foreland Basin in Bavaria (Germany)" (PDF). Newsletters on Stratigraphy. 49 (3): 445–467. doi:10.1127/nos/2016/0288.
- 1 2 3 Prieto, Jérôme; Ostende, Lars W. van den Hoek; Böhme, Madelaine; Braze, M. (2011). "Reappearance of Galerix (Erinaceomorpha, Mammalia) at the Middle to Late Miocene transition in South Germany: biostratigraphic and palaeoecologic implications" (PDF). Contributions to Zoology. 80 (3): 179. doi:10.1163/18759866-08003002. ISSN 1383-4517. S2CID 54798230.
- 1 2 3 4 5 Prieto, Jerome; Rummel, Michael (May 2009). "Evolution of the genus Collimys DAXNER-HÖCK, 1972 (Rodentia, Cricetidae) - A key to Middle to Late Miocene biostratigraphy in Central Europe". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 252/2 (2): 237–247. doi:10.1127/0077-7749/2009/0252-0237.
- 1 2 3 Kargopoulos, Nikolaos; Kampouridis, Panagiotis; Lechner, Thomas; Böhme, Madelaine (2022-11-02). "Hyaenidae (Carnivora) from the Late Miocene hominid locality of Hammerschmiede (Bavaria, Germany)" (PDF). Historical Biology. 34 (11): 2249–2258. doi:10.1080/08912963.2021.2010193. ISSN 0891-2963. S2CID 244913608.
- 1 2 3 4 5 6 7 8 Mayr, Gerald; Lechner, Thomas; Böhme, Madelaine (2020-05-06). "The large-sized darter Anhinga pannonica (Aves, Anhingidae) from the late Miocene hominid Hammerschmiede locality in Southern Germany". PLOS ONE. 15 (5): e0232179. Bibcode:2020PLoSO..1532179M. doi:10.1371/journal.pone.0232179. ISSN 1932-6203. PMC 7202596. PMID 32374733.
- 1 2 3 4 5 6 Schneider, Simon; Prieto, Jerome (2011-06-01). "First record of an autochthonous community of fluviatile freshwater molluscs from the Middle/Late Miocene Upper Freshwater Molasse (southern Germany)". Archiv für Molluskenkunde International Journal of Malacology. 140: 1–18. doi:10.1127/arch.moll/1869-0963/140/001-018.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 Kovar-Eder, Johanna (1984). "Pannonian (Upper Miocene) Vegetational Character and Climatic Inferences in the Central Paratethys Area". Annalen des Naturhistorischen Museums in Wien. Serie A für Mineralogie und Petrographie, Geologie und Paläontologie, Anthropologie und Prähistorie. 88: 117–129. ISSN 0255-0091. JSTOR 41701857.
- 1 2 3 4 5 6 Böhme, Madelaine; Ilg, August; Winklhofer, Michael (2008-11-15). "Late Miocene "washhouse" climate in Europe". Earth and Planetary Science Letters. 275 (3): 393–401. Bibcode:2008E&PSL.275..393B. doi:10.1016/j.epsl.2008.09.011. ISSN 0012-821X.
- ↑ Quan, Cheng; Liu, Yu-Sheng (Christopher); Tang, Hui; Utescher, Torsten (2014-07-11). "Miocene shift of European atmospheric circulation from trade wind to westerlies". Scientific Reports. 4 (1): 5660. Bibcode:2014NatSR...4E5660Q. doi:10.1038/srep05660. ISSN 2045-2322. PMC 4092332. PMID 25012454. S2CID 106499.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Kargopoulos, Nikolaos (2022-08-16). The carnivorans (Carnivora, Mammalia) from the hominid locality of Hammerschmiede (Bavaria, Germany) (Dissertation thesis). Universität Tübingen.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 Kargopoulos, Nikolaos; Valenciano, Alberto; Abella, Juan; Kampouridis, Panagiotis; Lechner, Thomas; Böhme, Madelaine (2022-07-13). "The exceptionally high diversity of small carnivorans from the Late Miocene hominid locality of Hammerschmiede (Bavaria, Germany)". PLOS ONE. 17 (7): e0268968. Bibcode:2022PLoSO..1768968K. doi:10.1371/journal.pone.0268968. ISSN 1932-6203. PMC 9278789. PMID 35830447.
- 1 2 Konidaris, George E.; Lechner, Thomas; Kampouridis, Panagiotis; Böhme, Madelaine (2023-12-01). "Deinotherium levius and Tetralophodon longirostris (Proboscidea, Mammalia) from the Late Miocene hominid locality Hammerschmiede (Bavaria, Germany), and their biostratigraphic significance for the terrestrial faunas of the European Miocene". Journal of Mammalian Evolution. 30 (4): 923–961. doi:10.1007/s10914-023-09683-3. ISSN 1573-7055.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 "Vertebrate Database fosFARbase: Amphibia, Birds, Fishes, Reptilia". www.wahre-staerke.com. Retrieved 2022-11-04.
- ↑ Gaudant, Jean (2015). "Re-examination of the upper Miocene freshwater fish fauna from Höwenegg (Hegau, Germany)". Swiss Journal of Palaeontology. 134 (1): 117–127. doi:10.1007/s13358-015-0073-2. ISSN 1664-2384. S2CID 84139965.
- ↑ Böhme, Madelaine (2002). "Freshwater fishes from the Pannonian of the Vienna Basin with special reference to the locality Sandberg near Götzendorf, Lower Austria" (PDF). Cour. Forsch.-Inst. Senckenberg. 237: 151–173.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 Klembara, Jozef; Böhme, Madelaine; Rummel, Michael (2010). "Revision of the anguine lizard Pseudopus laurillardi (Squamata, Anguidae) from the Miocene of Europe, with comments on paleoecology". Journal of Paleontology. 84 (2): 159–196. doi:10.1666/09-033R1.1. ISSN 0022-3360. JSTOR 40605510. S2CID 130738857.
- ↑ Schleich, Hans Hermann (20 December 1984). "Neue Reptilienfunde aus dem Tertiär Deutschlands. 1. Schildkröten aus dem Jungtertiär Süddeutschlands" (PDF). Naturwissenschaftliche Zeitschrift für Niederbayern.
- ↑ "Lottis Urahnen aus der Hammerschmiede: Im Allgäu lebten einst Alligatorschildkröten". Allgäuer Zeitung. 28 August 2022. Retrieved 2 November 2022.
- ↑ "Der größte Saurier aus der Hammerschmiede: Die Panzerschleiche". Allgäuer Zeitung. 1 May 2021. Archived from the original on 1 November 2022.
- 1 2 3 4 Mayr, Gerald; Lechner, Thomas; Böhme, Madelaine (2022-03-07). "Nearly complete leg of an unusual, shelduck-sized anseriform bird from the earliest late Miocene hominid locality Hammerschmiede (Germany)". Historical Biology. 35 (4): 465–474. doi:10.1080/08912963.2022.2045285. ISSN 0891-2963. S2CID 247310405.
- ↑ Mayr, Gerald; Lechner, Thomas; Böhme, Madelaine (2020-10-01). "A skull of a very large crane from the late Miocene of Southern Germany, with notes on the phylogenetic interrelationships of extant Gruinae". Journal of Ornithology. 161 (4): 923–933. doi:10.1007/s10336-020-01799-0. ISSN 2193-7206. S2CID 220505689.
- ↑ "Udos Kollege: Das Schwein mit dem Zungenbrecher-Namen". Allgäuer Zeitung. 19 December 2020. Retrieved 4 November 2022.
- ↑ Hartung, Josephina; Böhme, Madelaine (2022-05-18). "Unexpected cranial sexual dimorphism in the tragulid Dorcatherium naui based on material from the middle to late Miocene localities of Eppelsheim and Hammerschmiede (Germany)". PLOS ONE. 17 (5): e0267951. Bibcode:2022PLoSO..1767951H. doi:10.1371/journal.pone.0267951. ISSN 1932-6203. PMC 9116667. PMID 35584185.
- ↑ Hartung, Josephina; Lechner, Thomas; Böhme, Madelaine (2020-12-18). "New cranial material of Miotragocerus monacensis (Mammalia: Bovidae) from the late Miocene hominid locality Hammerschmiede (Germany)". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 298 (3): 269–284. doi:10.1127/njgpa/2020/0948. S2CID 229409794.
- ↑ Fuss, Jochen; Prieto, Jérôme; Böhme, Madelaine (2015-06-01). "Revision of the boselaphin bovid Miotragocerus monacensis Stromer, 1928 (Mammalia, Bovidae) at the Middle to Late Miocene transition in Central Europe". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 276 (3): 229–265. doi:10.1127/njgpa/2015/0481.
- ↑ Kargopoulos, Nikolaos; Valenciano, Alberto; Kampouridis, Panagiotis; Lechner, Thomas; Böhme, Madelaine (2021-05-04). "New early late Miocene species of Vishnuonyx (Carnivora, Lutrinae) from the hominid locality of Hammerschmiede, Bavaria, Germany". Journal of Vertebrate Paleontology. 41 (3): e1948858. doi:10.1080/02724634.2021.1948858. ISSN 0272-4634. S2CID 240538139.
- ↑ Kargopoulos, Nikolaos; Kampouridis, Panagiotis; Lechner, Thomas; Böhme, Madelaine (2021-12-01). "A review of Semigenetta (Viverridae, Carnivora) from the Miocene of Eurasia based on material from the hominid locality of Hammerschmiede (Germany)". Geobios. 69: 25–36. doi:10.1016/j.geobios.2021.07.001. ISSN 0016-6995. S2CID 237805066.
- ↑ Prieto, Jérôme; van Dam, Jan A. (2012-11-01). "Primitive Anourosoricini and Allosoricinae from the Miocene of Germany". Geobios. 45 (6): 581–589. doi:10.1016/j.geobios.2012.03.001. ISSN 0016-6995.
- ↑ van Dam, Jan A. (2010-07-14). "The systematic position of Anourosoricini (Soricidae, Mammalia): paleontological and molecular evidence". Journal of Vertebrate Paleontology. 30 (4): 1221–1228. doi:10.1080/02724634.2010.483553. ISSN 0272-4634. S2CID 220413165.
- ↑ Scott A. Williams et al.: Reevaluating bipedalism in Danuvius. In: Nature. Band 586, 2020, S. E1–E3, doi:10.1038/s41586-020-2736-4. Madelaine Böhme et al.: Reply to: Reevaluating bipedalism in Danuvius. In: Nature. Band 586, 2020, S. E4–E5, doi:10.1038/s41586-020-2737-3. Fossils suggest tree-dwelling apes walked upright long before hominids did. Archived 2022-11-11 at the Wayback Machine Auf: sciencenews.org vom 6. November 2019.
- ↑ "Raufbolde und Einhörner: Vor Millionen Jahren lebten im Allgäu Nashörner". Allgäuer Zeitung. 18 April 2021. Retrieved 4 November 2022.
- ↑ "So ähnlich sah das größte Landsäugetier aus dem Allgäu aus". Allgäuer Zeitung. 18 July 2020. Retrieved 4 November 2022.
- ↑ Prieto, Jérome (2007-12-20). Kleinsäuger-Biostratigraphie und Paläoökologie des höheren Mittelmiozäns (MN 8) Bayerns (Text.PhDThesis thesis) (in German). Ludwig-Maximilians-Universität München.
- ↑ Casanovas-Vilar, Isaac; Alba, David M.; Almécija, Sergio; Robles, Josep M.; Galindo, Jordi; Moyà-Solà, Salvador (2008). "Taxonomy and Paleobiology of the Genus Chalicomys Kaup, 1832 (Rodentia, Castoridae), with the Description of a New Species from Abocador De Can Mata (Vallès-Penedès Basin, Catalonia, Spain)". Journal of Vertebrate Paleontology. 28 (3): 851–862. doi:10.1671/0272-4634(2008)28[851:TAPOTG]2.0.CO;2. ISSN 0272-4634. JSTOR 20491008. S2CID 85823651.
- ↑ "The beaver Steneofiber depereti from the lower Upper Miocene hominid locality Hammerschmiede and remarks on its ecology - Acta Palaeontologica Polonica". app.pan.pl. Retrieved 2022-11-24.
- ↑ Daxner-Höck, Gudrun (January 2004). "Biber und ein Zwerghamster aus Mataschen (Unter-Pannonium, Steirisches Becken)". Joannea – Geologie und Paläontologie.
- ↑ García-Paredes, Israel; Peláez-Camponanes, Pablo; Álvarez-Sierra, M. Ángeles (2010-09-01). "Microdyromys remmerti, sp. nov., a new Gliridae (Rodentia, Mammalia) from the Aragonian type area (Miocene, Calatayud-Montalbán basin, Spain)". Journal of Vertebrate Paleontology. 30 (5): 1594–1609. doi:10.1080/02724634.2010.501453. ISSN 0272-4634. S2CID 84520113.
- ↑ Prieto, Jérôme (January 2012). "The Genus Eomyops Engesser, 1979 (Rodentia, Eomyidae) from the youngest deposits of the German part of the North Alpine Foreland Basin". Swiss Journal of Palaeontology. 131 (1): 95–106. doi:10.1007/s13358-011-0033-4. ISSN 1664-2384. S2CID 129383145.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 Seitner, Liselotte (2004-09-23). "Miocene microflora of Hammerschmiede (Germany)". In supplement to: Seitner, L (1987): Miozäne Mikrofloren aus Sedimenten der Süssbrackwassermolasse und der Oberen Süsswassermolasse Süddeutschlands. PhD Thesis, Fakultät für Geowissenschaften der Ludwig-Maximilians-Universität München, Germany, 1-352. doi:10.1594/PANGAEA.206523.
- ↑ Fischer, Thilo C.; Butzmann, Rainer; Meller, Barbara; Rattei, Thomas; Newman, Mark; Hölscher, Dirk (2009). "The morphology, systematic position and inferred biology of Spirematospermum — An extinct genus of Zingiberales". Review of Palaeobotany and Palynology. 157 (3–4): 391–426. doi:10.1016/j.revpalbo.2009.06.010. ISSN 0034-6667.