| Ventrolateral Preoptic Nucleus | |
|---|---|
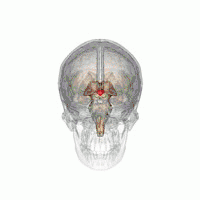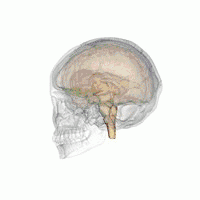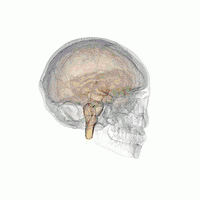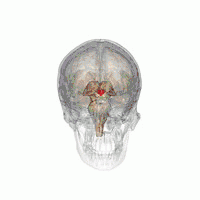 The VLPO is located at the anterior of the hypothalamus. It is also called the intermediate nucleus of the preoptic area | |
| Details | |
| Part of | Preoptic nucleus |
| Identifiers | |
| Acronym(s) | VLPO or IPA |
| NeuroNames | 3122 |
| Anatomical terms of neuroanatomy | |
The ventrolateral preoptic nucleus (VLPO), also known as the intermediate nucleus of the preoptic area (IPA), is a small cluster of neurons situated in the anterior hypothalamus, sitting just above and to the side of the optic chiasm in the brain of humans and other animals.[1][2] The brain's sleep-promoting nuclei (e.g., the VLPO, parafacial zone, nucleus accumbens core, and lateral hypothalamic MCH neurons),[3][4][5][6] together with the ascending arousal system which includes components in the brainstem, hypothalamus and basal forebrain, are the interconnected neural systems which control states of arousal, sleep, and transitions between these two states.[1][4][5] The VLPO is active during sleep, particularly during non-rapid eye movement sleep (NREM sleep),[7] and releases inhibitory neurotransmitters, mainly GABA and galanin, which inhibit neurons of the ascending arousal system that are involved in wakefulness and arousal.[1][8] The VLPO is in turn innervated by neurons from several components of the ascending arousal system.[9] The VLPO is activated by the endogenous sleep-promoting substances adenosine[10][11] and prostaglandin D2.[12] The VLPO is inhibited during wakefulness by the arousal-inducing neurotransmitters norepinephrine and acetylcholine.[9] The role of the VLPO in sleep and wakefulness, and its association with sleep disorders – particularly insomnia and narcolepsy – is a growing area of neuroscience research.
Structure
At least 80% of neurons in the VLPO that project to the ascending arousal system are GABAergic (neurons that produce GABA).[13] In vitro studies in rats have shown that many neurons in the VLPO that are inhibited by norepinephrine or acetylcholine are multipolar triangular shaped cells with low threshold spikes.[9] These triangular multipolar neurons exist in two sub-populations in the VLPO:
- Type 1 – inhibited by serotonin.
- Type 2 – excited by serotonin and adenosine.
As adenosine accumulates during wakefulness[11] it is likely that type 2 cells play a role in sleep induction.
The remaining third of neurons in the VLPO are excited by norepinephrine. Their role is unclear.
Function
Sleep/wakefulness
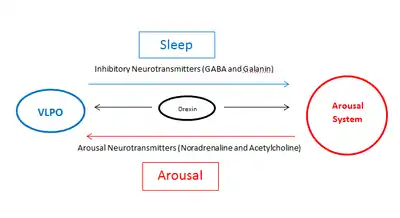
In the early 20th century, Constantin von Economo noted that humans who had encephalitis with lesions in the anterior hypothalamus had insomnia, and proposed a sleep-promoting influence from that area.[2] Animal studies in the mid-20th century in rats and cats confirmed that very large lesions in the preoptic area and basal forebrain resulted in insomnia[14][15] but did not identify the cell group that was responsible. In 1996, Sherin and colleagues reported the presence of a cell group in the VLPO that expresses cFos (a protein often found in neurons that have recently been active) during sleep, and that these neurons contain the inhibitory neurotransmitters GABA and galanin.[13][7] These same neurons were found to innervate components of the ascending arousal system, including the tuberomammillary nucleus (TMN) and other components of the lateral hypothalamus; the raphe nuclei; the locus coeruleus (LC); the pedunculopontine (PPT) and laterodorsal tegmental nuclei (LDT); and the parabrachial nucleus (PB). More recent studies using opto- or chemogenetic activation of VLPO neurons have confirmed that they promote sleep.[16]
The sleep-promoting effects of the VLPO neurons is thought to be due to release of GABA and possibly galanin that suppresses firing of arousal system neurons. As the VLPO is also inhibited by neurotransmitters released by components of the arousal systems, such as acetylcholine and norepinephrine,[9] a current theory has proposed that the VLPO and the arousal system form a "flip-flop" circuit.[1][2] This term from electrical engineering denotes a circuit in which mutual inhibition means that each component of the circuit, as it turns on, turns the other off, resulting in rapid transitions from one state (wake or sleep) to the other, with minimal time in transition states. This theory has been used to create mathematical models that explain much of the wake-sleep behavior in animals, including in pathological states and responses to drugs.[1][17][18] Orexin neurons in the posterior lateral hypothalamus potentiate neurons in the ascending arousal system and help stabilize the brain in the waking state (and consolidated wakefulness, which builds up homeostatic sleep drive, helps stabilize the brain during later sleep). The loss of orexin neurons in the disorder narcolepsy destabilizes the wake-sleep switch, resulting in overwhelming sleep episodes during the waking day, as well as more frequent awakenings from sleep at night.[1]
Circadian rhythm
There is a strong circadian rhythm of sleep in mammals. The “master clock” for circadian rhythms in mammals is the suprachiasmatic nucleus (SCN). The SCN has little if any projection directly to the VLPO neurons. Instead, they project strongly to the adjacent subparaventricular zone, which in turn contains inhibitory GABAergic neurons that innervate the dorsomedial nucleus of the hypothalamus.[19][20] Lesions of the dorsomedial nucleus almost completely eliminate the circadian rhythm of sleep. GABAergic neurons in the dorsomedial nucleus innervate the VLPO, and glutamatergic neurons innervate the lateral hypothalamus, suggesting that the dorsomedial nucleus mainly promotes wakefulness during the active period (daytime for humans).[19]
Clinical significance
Insomnia
Elderly human patients with more galanin neurons in their intermediate nucleus (the human equivalent of the VLPO galanin neurons in rodents) have better, more continuous sleep. A reduced number of VLPO neurons is associated with more fragmented sleep (more awakenings throughout the night).[21]
Lesions in the VLPO in rats results in 50-60% decrease in NREM sleep time and prolonged insomnia.[22] More recent research suggests that stress-induced insomnia could be due to an imbalance of input to arousal system and VLPO neurons.[23]
Sedative/hypnotic drugs
Many sedative/hypnotic drugs act by binding to and potentiating GABA-A receptors. These include older drugs such as ethanol, chloral hydrate and barbiturates, as well as newer benzodiazepines and "non-benzodiazepine" drugs (such as zolpidem, which bind to the same receptor but have a different chemical configuration), and even anesthetics such as propofol and isoflurane. As the VLPO inputs to the arousal system use this same receptor, these drugs at low doses essentially act by potentiating the VLPO, producing a sleepy state. Animal studies show that VLPO neurons show cFos activation after sedative doses of these drugs,[24] and that VLPO lesions produce resistance to their sedative effects.[25] However, at high doses that produce a surgical plane of anesthesia, these drugs have much more widespread inhibitory effects, that do not depend upon the VLPO.[26] Studies have shown that multiple sedative/hypnotic drugs that act by potentiating GABA-A receptors, including ethanol, chloral hydrate, propofol and gas anesthetics such as isoflurane, at sedative doses increase the activity of the VLPO neurons in mice.[27] This finding suggests that at relatively low sedative doses, these medications may have a common mechanism of action, which includes potentiating the firing of VLPO neurons. High doses used in surgical anesthesia, however, reduce activity of neurons throughout the nervous system.
References
- 1 2 3 4 5 6 Saper, Clifford B.; Fuller, Patrick M.; Pedersen, Nigel P.; Lu, Jun; Scammell, Thomas E. (2010). "Sleep State Switching". Neuron. 68 (6): 1023–1042. doi:10.1016/j.neuron.2010.11.032. PMC 3026325. PMID 21172606.
- 1 2 3 Saper, Clifford B.; Scammell, Thomas E.; Lu, Jun (2005). "Hypothalamic regulation of sleep and circadian rhythms". Nature. 437 (7063): 1257–1263. doi:10.1038/nature04284. ISSN 0028-0836. PMID 16251950. S2CID 1793658.
- ↑ Anaclet C, Ferrari L, Arrigoni E, Bass CE, Saper CB, Lu J, Fuller PM (September 2014). "The GABAergic parafacial zone is a medullary slow wave sleep-promoting center" (PDF). Nat. Neurosci. 17 (9): 1217–1224. doi:10.1038/nn.3789. PMC 4214681. PMID 25129078.
In the present study we show, for the first time, that activation of a delimited node of GABAergic neurons located in the medullary PZ can potently initiate SWS and cortical SWA in behaving animals. ... For now however it remains unclear if the PZ is interconnected with other sleep– and wake–promoting nodes beyond the wake–promoting PB. ... The intensity of cortical slow–wave–activity (SWA: 0.5–4Hz) during SWS is also widely accepted as a reliable indicator of sleep need ... In conclusion, in the present study we demonstrated that all polygraphic and neurobehavioral manifestation of SWS, including SWA, can be initiated in behaving animals by the selective activation of a delimited node of GABAergic medullary neurons.
- 1 2 Schwartz MD, Kilduff TS (December 2015). "The Neurobiology of Sleep and Wakefulness". The Psychiatric Clinics of North America. 38 (4): 615–644. doi:10.1016/j.psc.2015.07.002. PMC 4660253. PMID 26600100.
More recently, the medullary parafacial zone (PZ) adjacent to the facial nerve was identified as a sleep-promoting center on the basis of anatomical, electrophysiological and chemo- and optogenetic studies.23, 24 GABAergic PZ neurons inhibit glutamatergic parabrachial (PB) neurons that project to the BF,25 thereby promoting NREM sleep at the expense of wakefulness and REM sleep. ... Sleep is regulated by GABAergic populations in both the preoptic area and the brainstem; increasing evidence suggests a role for the melanin-concentrating hormone cells of the lateral hypothalamus and the parafacial zone of the brainstem
- 1 2 Brown RE, McKenna JT (June 2015). "Turning a Negative into a Positive: Ascending GABAergic Control of Cortical Activation and Arousal". Front. Neurol. 6: 135. doi:10.3389/fneur.2015.00135. PMC 4463930. PMID 26124745.
The sleep-promoting action of GABAergic neurons located in the preoptic hypothalamus (6–8) is now well-known and accepted (9). More recently, other groups of sleep-promoting GABAergic neurons in the lateral hypothalamus (melanin-concentrating hormone neurons) and brainstem [parafacial zone; (10)] have been identified.
- ↑ Oishi Y, Xu Q, Wang L, Zhang BJ, Takahashi K, Takata Y, Luo YJ, Cherasse Y, Schiffmann SN, de Kerchove d'Exaerde A, Urade Y, Qu WM, Huang ZL, Lazarus M (September 2017). "Slow-wave sleep is controlled by a subset of nucleus accumbens core neurons in mice". Nature Communications. 8 (1): 734. doi:10.1038/s41467-017-00781-4. PMC 5622037. PMID 28963505.
Here, we show that chemogenetic or optogenetic activation of excitatory adenosine A2A receptor-expressing indirect pathway neurons in the core region of the NAc strongly induces slow-wave sleep. Chemogenetic inhibition of the NAc indirect pathway neurons prevents the sleep induction, but does not affect the homoeostatic sleep rebound.
- 1 2 Sherin, J. E.; Shiromani, P. J.; McCarley, R. W.; Saper, C. B. (1996-01-12). "Activation of Ventrolateral Preoptic Neurons During Sleep". Science. 271 (5246): 216–219. doi:10.1126/science.271.5246.216. ISSN 0036-8075. PMID 8539624. S2CID 19804653.
- ↑ Brown, Ritchie E.; Basheer, Radhika; McKenna, James T.; Strecker, Robert E.; McCarley, Robert W. (2012). "Control of Sleep and Wakefulness". Physiological Reviews. 92 (3): 1087–1187. doi:10.1152/physrev.00032.2011. ISSN 0031-9333. PMC 3621793. PMID 22811426.
- 1 2 3 4 Gallopin, Thierry; Fort, Patrice; Eggermann, Emmanuel; Cauli, Bruno; Luppi, Pierre-Hervé; Rossier, Jean; Audinat, Etienne; Mühlethaler, Michel; Serafin, Mauro (2000). "Identification of sleep-promoting neurons in vitro". Nature. 404 (6781): 992–995. doi:10.1038/35010109. ISSN 0028-0836. PMID 10801127. S2CID 4364527.
- ↑ Scammell, T.E; Gerashchenko, D.Y; Mochizuki, T; McCarthy, M.T; Estabrooke, I.V; Sears, C.A; Saper, C.B; Urade, Y; Hayaishi, O (2001). "An adenosine A2a agonist increases sleep and induces Fos in ventrolateral preoptic neurons". Neuroscience. 107 (4): 653–663. doi:10.1016/s0306-4522(01)00383-9. ISSN 0306-4522. PMID 11720788. S2CID 45189308.
- 1 2 Gallopin T (2005). "The endogenous somnogen adenosine excites a subset of sleep-promoting neurons via A2A receptors in the ventrolateral preoptic nucleus". Neuroscience. 134 (4): 1377–1390. doi:10.1016/j.neuroscience.2005.05.045. PMID 16039802. S2CID 27732204.
- ↑ Scammell T (1998). "Activation of ventrolateral preoptic neurons by the somnogen prostaglandin D2". PNAS. 95 (13): 7754–7759. doi:10.1073/pnas.95.13.7754. PMC 22747. PMID 9636223.
- 1 2 Sherin J (1998). "Innervation of Histaminergic Tuberomammillary Neurons by GABAergic and Galaninergic Neurons in the Ventrolateral Preoptic Nucleus of the Rat". The Journal of Neuroscience. 18 (12): 4705–4721. doi:10.1523/JNEUROSCI.18-12-04705.1998. PMC 6792696. PMID 9614245.
- ↑ Nauta W (1946). "Hypothalamic regulation of sleep in rats". Journal of Neurophysiology. 9: 285–314. doi:10.1152/jn.1946.9.4.285. PMID 20991815.
- ↑ McGinty D (1968). "Sleep Suppression after Basal Forebrain Lesions in the Cat". Science. 160 (3833): 1253–1255. doi:10.1126/science.160.3833.1253. PMID 5689683. S2CID 24677928.
- ↑ Kroeger, Daniel; Absi, Gianna; Gagliardi, Celia; Bandaru, Sathyajit S.; Madara, Joseph C.; Ferrari, Loris L.; Arrigoni, Elda; Münzberg, Heike; Scammell, Thomas E. (2018). "Galanin neurons in the ventrolateral preoptic area promote sleep and heat loss in mice". Nature Communications. 9 (1): 4129. doi:10.1038/s41467-018-06590-7. ISSN 2041-1723. PMC 6175893. PMID 30297727.
- ↑ Phillips, A.J.K.; Robinson, P.A. (2007). "A Quantitative Model of Sleep-Wake Dynamics Based on the Physiology of the Brainstem Ascending Arousal System". Journal of Biological Rhythms. 22 (2): 167–179. doi:10.1177/0748730406297512. ISSN 0748-7304. PMID 17440218. S2CID 31817324.
- ↑ Fulcher, Ben D.; Phillips, Andrew J. K.; Postnova, Svetlana; Robinson, Peter A. (2014). Mistlberger, Ralph E. (ed.). "A Physiologically Based Model of Orexinergic Stabilization of Sleep and Wake". PLOS ONE. 9 (3): e91982. doi:10.1371/journal.pone.0091982. ISSN 1932-6203. PMC 3961294. PMID 24651580.
- 1 2 Chou, Thomas C.; Scammell, Thomas E.; Gooley, Joshua J.; Gaus, Stephanie E.; Saper, Clifford B.; Lu, Jun (2003). "Critical Role of Dorsomedial Hypothalamic Nucleus in a Wide Range of Behavioral Circadian Rhythms". The Journal of Neuroscience. 23 (33): 10691–10702. doi:10.1523/jneurosci.23-33-10691.2003. ISSN 0270-6474. PMC 6740926. PMID 14627654.
- ↑ Vujovic, Nina; Gooley, Joshua J.; Jhou, Thomas C.; Saper, Clifford B. (2015). "Projections from the subparaventricular zone define four channels of output from the circadian timing system: Projections from the SPZ". Journal of Comparative Neurology. 523 (18): 2714–2737. doi:10.1002/cne.23812. PMC 4607558. PMID 26010698.
- ↑ Lim A (2014). "Sleep is related to neuron numbers in the ventrolateral preoptic/intermediate nucleus in older adults with and without Alzheimer's disease". Brain. 137 (Pt 10): 2847–61. doi:10.1093/brain/awu222. PMC 4163039. PMID 25142380.
- ↑ Lu, Jun; Greco, Mary Ann; Shiromani, Priyattam; Saper, Clifford B. (2000). "Effect of Lesions of the Ventrolateral Preoptic Nucleus on NREM and REM Sleep". The Journal of Neuroscience. 20 (10): 3830–3842. doi:10.1523/jneurosci.20-10-03830.2000. ISSN 0270-6474. PMC 6772663. PMID 10804223.
- ↑ Cano, G.; Mochizuki, T.; Saper, C. B. (2008). "Neural Circuitry of Stress-Induced Insomnia in Rats". Journal of Neuroscience. 28 (40): 10167–10184. doi:10.1523/JNEUROSCI.1809-08.2008. ISSN 0270-6474. PMC 2693213. PMID 18829974.
- ↑ Lu, Jun; Nelson, Laura E.; Franks, Nick; Maze, Mervyn; Chamberlin, Nancy L.; Saper, Clifford B. (2008). "Role of endogenous sleep-wake and analgesic systems in anesthesia". The Journal of Comparative Neurology. 508 (4): 648–662. doi:10.1002/cne.21685. PMC 4924624. PMID 18383504.
- ↑ Moore, Jason T.; Chen, Jingqiu; Han, Bo; Meng, Qing Cheng; Veasey, Sigrid C.; Beck, Sheryl G.; Kelz, Max B. (2012). "Direct Activation of Sleep-Promoting VLPO Neurons by Volatile Anesthetics Contributes to Anesthetic Hypnosis". Current Biology. 22 (21): 2008–2016. doi:10.1016/j.cub.2012.08.042. PMC 3628836. PMID 23103189.
- ↑ Eikermann, Matthias; Vetrivelan, Ramalingam; Grosse-Sundrup, Martina; Henry, Mark E.; Hoffmann, Ulrike; Yokota, Shigefumi; Saper, Clifford B.; Chamberlin, Nancy L. (2011). "The ventrolateral preoptic nucleus is not required for isoflurane general anesthesia". Brain Research. 1426: 30–37. doi:10.1016/j.brainres.2011.10.018. PMC 3215917. PMID 22041226.
- ↑ Moore J (2012). "Direct Activation of Sleep-Promoting VLPO Neurons by Volatile Anesthetics Contributes to Anesthetic Hypnosis". Current Biology. 22 (21): 2008–2016. doi:10.1016/j.cub.2012.08.042. PMC 3628836. PMID 23103189.
External links
Gallopin T, Luppi PH, Cauli B, Urade Y, Rossier J, Hayaishi O, Lambolez B, Fort P (2005). "The endogenous somnogen adenosine excites a subset of sleep-promoting neurons via A2A receptors in the ventrolateral preoptic nucleus". Neuroscience. 134 (4): 1377–90. doi:10.1016/j.neuroscience.2005.05.045. PMID 16039802. S2CID 27732204.