| Marsupial moles[1] Temporal range: Miocene to Recent | |
|---|---|
_(cropped).jpg.webp) | |
| Southern marsupial mole | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Infraclass: | Marsupialia |
| Order: | Notoryctemorphia |
| Family: | Notoryctidae |
| Genus: | Notoryctes Stirling, 1891 |
| Type species | |
| Psammoryctes typhlops Stirling, 1889 | |
| Species | |
|
Notoryctes typhlops Stirling, 1891 | |
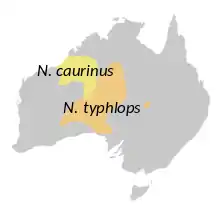 | |
| Ranges of the two species | |
Marsupial moles, the Notoryctidae /noʊtəˈrɪktɪdiː/ family, are two species of highly specialized marsupial mammals that are found in the Australian interior.[2] They are small fossorial marsupials that anatomically converge on fossorial placental mammals, such as extant golden moles (Chrysochloridae) and extinct epoicotheres (Pholidota). The species are:
- Notoryctes typhlops (southern marsupial mole, known as the itjaritjari by the Pitjantjatjara and Yankunytjatjara people in Central Australia)[3]
- Notoryctes caurinus (northern marsupial mole, also known as the kakarratul)
Characteristics
In an example of convergent evolution, notoryctids resemble (and fill the ecological niche of) the talpid or true moles from North America and Eurasia and the Chrysochloridae or golden moles from Southern Africa. Like chrysochlorids and epoicotheres, notoryctids use their forelimbs and enlarged central claws to dig in a parasagittal (i.e., up and down) plane, as opposed to the "lateral scratch" style of digging that characterizes talpid moles.[4][5]
Marsupial moles spend most of their time underground, coming to the surface only occasionally, probably mostly after rains. They are functionally blind, their eyes having become reduced to vestigial lenses under the skin that lack a pupil. They have no external ears, just a pair of tiny holes hidden under thick hair. The head is cone-shaped, with a leathery shield over the muzzle, the body is tubular, and the tail is a short, bald stub encased in leathery skin. They are between 12 and 16 centimetres (4.7–6.3 in) long, weigh 40 to 60 grams (1.4–2.1 oz), and are uniformly covered in fairly short, very fine pale cream to white hair with an iridescent golden sheen. Their pouch is small but well developed and has evolved to face backwards so it does not fill with sand. It contains just two teats, so the animal cannot support more than two young at a time.
The limbs are very short, with reduced digits. The forefeet have two greatly enlarged, spade shaped, flat claws on the third and fourth digits, which are used to excavate soil in front of the animal. The hindfeet are flattened, and bear three small claws; these feet are used to push soil behind the animal as it digs. Epipubic bones are present but small and as in some other fossorial mammals (e.g., armadillos), the last five cervical vertebrae are fused to give the head greater rigidity during digging. The animal swims through the soil and does not leave behind a permanent burrow.
The teeth of the marsupial moles are degenerate and bear no resemblance to polyprotodont or diprotodont teeth. Their dental formula varies, but is usually somewhere near 4–3.1.2.43.1.3.4 × 2 = 42–44. The upper molar teeth are triangular and zalambdodont, i.e., resembling an inverted Greek letter lambda in occlusal view, and the lower molars appear to have lost their talonid basins.[6][7][8]
Fossil record
Notoryctids are represented by early Miocene fossils of Naraboryctes from Riversleigh in Queensland, Australia, which document the mosaic acquisition of dental and skeletal features of the living Notoryctes from a more surface-dwelling ancestor.[9] The notoryctid fossil record demonstrates that the primary cusp of the molars is the metacone,[9] distinct from the paracone characteristic of zalambdodont tenrecs, golden moles, and Solenodon.[10] Regarding the number of teeth in each dental quadrant (or dental formula) Archer et al. (2011) reported that "the dental formula for species of Notoryctes is controversial because of considerable polymorphism in tooth number, both between specimens and within the same specimen." Nonetheless, reflecting the consensus of older studies,[11] Archer et al. (2011) report the presence of four molars (typical for marsupials) in each quadrant both in living Notoryctes and the fossil notoryctid Naraboryctes.
Evolutionary affinities
American paleontologist William King Gregory wrote in 1910 (p. 209) that "Notoryctes is a true marsupial" and this view has been repeatedly verified by phylogenetic analyses of comparative anatomy,[12][13] mitochondrial DNA,[14][15][16] nuclear DNA,[17][18] rare genomic events,[19] and combined datasets of nuclear and mitochondrial DNA[20] and morphology and DNA.[21][22][23] The largest phylogenetic datasets strongly support the placement of Notoryctes as the sister taxon to a dasyuromorph-peramelian clade, within the Australidelphian radiation.[24]
References
- ↑ Groves, C. P. (2005). Wilson, D. E.; Reeder, D. M. (eds.). Mammal Species of the World: A Taxonomic and Geographic Reference (3rd ed.). Baltimore: Johns Hopkins University Press. p. 22. ISBN 0-801-88221-4. OCLC 62265494.
- ↑ Warburton N. Functional morphology and evolution of marsupial moles (Marsupialia: Notoryctemorphia). 2003. PhD Dissertation, University of Western Australia.
- ↑ "Mole Patrol". The Marsupial Society. 2004. Archived from the original on 2008-02-04. Retrieved 2006-11-09.
- ↑ Yalden, D.W. (1 May 1966). "The anatomy of mole locomotion". Journal of Zoology. 149 (1): 55–64. doi:10.1111/j.1469-7998.1966.tb02983.x.
- ↑ Rose KD, Emry RJ (1 January 1983). "Extraordinary fossorial adaptations in the Oligocene palaeanodonts Epoicotherium and Xenocranium (Mammalia)". Journal of Morphology. 175 (1): 33–56. doi:10.1002/jmor.1051750105. PMID 30053775. S2CID 51727274.
- ↑ "Notoryctes caurinus". animaldiversity.org.
- ↑ Asher RJ, Horovitz I, Martin T, Sanchez-Villagra MR (15 January 2007). "Neither a rodent nor a platypus: a reexamination of Necrolestes patagonensis Ameghino". American Museum Novitates (3546): 1–40. doi:10.1206/0003-0082(2007)3546[1:NARNAP]2.0.CO;2. S2CID 83642532.
- ↑ Gordon, Greg (1984). Macdonald, D. (ed.). The Encyclopedia of Mammals. New York: Facts on File. p. 842. ISBN 978-0-87196-871-5.
- 1 2 Archer, M. (2011-05-22). "Australia's first fossil marsupial mole (Notoryctemorphia) resolves controversies about their evolution and palaeoenvironmental origins". Proceedings of the Royal Society of London B: Biological Sciences. 278 (1711): 1498–1506. doi:10.1098/rspb.2010.1943. PMC 3081751. PMID 21047857.
- ↑ Asher RJ, Sánchez-Villagra MR. Locking yourself out: diversity among dentally zalambdodont therian mammals. Journal of Mammalian Evolution. 2005 Jun 1;12(1):265-82.
- ↑ Thomas O. XI.—Notoryctes in North-west Australia. Journal of Natural History. 1920 Jul 1;6(31):111-3.
- ↑ Horovitz I, Sánchez-Villagra MR. A morphological analysis of marsupial mammal higher‐level phylogenetic relationships. Cladistics. 2003 Jun 1;19(3):181-212.
- ↑ Beck RM, Warburton NM, Archer MI, Hand SJ, Aplin KP. Going underground: postcranial morphology of the early Miocene marsupial mole Naraboryctes philcreaseri and the evolution of fossoriality in notoryctemorphians. Memoirs of Museum Victoria. 2016;74:151-71.
- ↑ Springer MS, Westerman M, Kavanagh JR, Burk A, Woodburne MO, Kao DJ, Krajewski C. The origin of the Australasian marsupial fauna and the phylogenetic affinities of the enigmatic monito del monte and marsupial mole. Proceedings of the Royal Society of London B: Biological Sciences. 1998 Dec 22;265(1413):2381-6.
- ↑ Burk A, Westerman M, Kao DJ, Kavanagh JR, Springer MS. An analysis of marsupial interordinal relationships based on 12S rRNA, tRNA valine, 16S rRNA, and cytochrome b sequences. Journal of Mammalian Evolution. 1999 Dec 1;6(4):317-34.
- ↑ Kjer KM, Honeycutt RL. Site specific rates of mitochondrial genomes and the phylogeny of eutheria. BMC Evolutionary Biology. 2007 Jan 25;7(1):8.
- ↑ Amrine-Madsen H, Scally M, Westerman M, Stanhope MJ, Krajewski C, Springer MS. Nuclear gene sequences provide evidence for the monophyly of australidelphian marsupials. Molecular Phylogenetics and Evolution. 2003 Aug 31;28(2):186-96.
- ↑ Meredith RW, Janečka JE, Gatesy J, Ryder OA, Fisher CA, Teeling EC, Goodbla A, Eizirik E, Simão TL, Stadler T, Rabosky DL. Impacts of the Cretaceous Terrestrial Revolution and KPg extinction on mammal diversification. Science. 2011 Oct 28;334(6055):521-4.
- ↑ Nilsson, M. A.; Churakov, G.; Sommer, M.; Van Tran, N.; Zemann, A.; Brosius, J.; Schmitz, J. (2010). "Tracking Marsupial Evolution Using Archaic Genomic Retroposon Insertions". PLOS Biology. Public Library of Science. 8 (7): e1000436. doi:10.1371/journal.pbio.1000436. PMC 2910653. PMID 20668664.
- ↑ Phillips MJ, McLenachan PA, Down C, Gibb GC, Penny D. Combined mitochondrial and nuclear DNA sequences resolve the interrelations of the major Australasian marsupial radiations. Systematic Biology. 2006 Feb 1;55(1):122-37.
- ↑ Asher RJ, Horovitz I, Sánchez-Villagra MR. 2004. First Combined Cladistic Analysis of Marsupial Mammal Interrelationships. Molecular Phylogenetics and Evolution 33: 240-250.
- ↑ Beck RM, Godthelp H, Weisbecker V, Archer M, Hand SJ. Australia's oldest marsupial fossils and their biogeographical implications. PLos one. 2008 Mar 26;3(3):e1858.
- ↑ O’Meara RN, Thompson RS. Were there Miocene meridiolestidans? Assessing the phylogenetic placement of Necrolestes patagonensis and the presence of a 40 million year meridiolestidan ghost lineage. Journal of Mammalian Evolution. 2014 Sep 1;21(3):271-84.
- ↑ Mitchell KJ, Pratt RC, Watson LN, Gibb GC, Llamas B, Kasper M, Edson J, Hopwood B, Male D, Armstrong KN, Meyer M. Molecular phylogeny, biogeography, and habitat preference evolution of marsupials. Molecular biology and evolution. 2014 Sep 1;31(9):2322-30.
- University of Western Australia marsupial mole home page
- Research mission to discover conservation requirements Archived 2005-09-03 at the Wayback Machine (includes photo)
- Nature news story (includes photo)
- AusEmade: Marsupial Mole Information
- Archer, Hand & Godthelp, Australia's lost world: Riversleigh, World Heritage Site, Reed New Holland, 1991. ISBN 1-876334-59-2
- Notoryctes caurinus, Animal Diversity Web
External links
- ARKive - images and movies of the marsupial mole (Notoryctes typhlops)
- Australian Geographic - The marsupial mole: an enduring enigma
| ||||||||||||||||||||||||||||||||||||||||
| 
 | |||||||||||||||||||||||||||||||||||||||
