

Perilipin 5, also known as Oxpatperilipin 5 or PLIN5, is a protein that belongs to perilipin family. This protein group has been shown to be responsible for lipid droplet's biogenesis, structure and degradation.[5] In particular, Perilipin 5 is a lipid droplet-associated protein whose function is to keep the balance between lipolysis and lipogenesis, as well as maintaining lipid droplet homeostasis. For example, in oxidative tissues, muscular tissues and cardiac tissues, PLIN5 promotes association between lipid droplets and mitochondria.[6]
Inside the cell, PLIN5 can be found in multiple intracellular structures including lipid droplets, endoplasmic reticulum, mitochondria, and the cytosol.[7] The wrong expression of this protein has been proven to be related with diseases such as skeletal muscle diseases, liver diseases or carcinogenesis.[8]
Perilipin Protein Family
PLIN5 is the fifth of the 5 perilipins which can be found on humans. All of them (PLIN1, ADRP, TIP47, S3-12 and PLIN5) have similar functionality, relating to lipid droplets. They have a 1,252% similarity with 76 identical positions. On the other hand, PLIN5 has its most similarity with PLINS 2 and 3, with over 150 similar positions and 18,644% similarity.
Structure
Perilipin 5 is a relatively large protein. It is composed of 463 amino acids, weighing an average of 50.8 kDa. It contains four basic regions:
- from amino acid 1 to 173: key region for lipid droplet targeting.
- from amino acid 1 to 108: interaction with LIPE, a gene that encodes the formation of hormone-sensitive lipase, also known as HSL, whose main function is to mobilize the previously stored fats.
- from amino acid 185 to 463: interactions with PNPLA2 and ABHD5.
- from amino acid 444 to 463: targets mitochondria for lipid droplet-mitochondria association.
PLIN5 is expressed in 183 organs, having its highest expression level on the stomach fundus.
Protein Kinase A (PKA) phosphorylates residues 2, 148 and 322. Phosphorylation by PKA enables lipolysis probably by promoting the release of ABHD5 from the perilipin scaffold.
Evolution
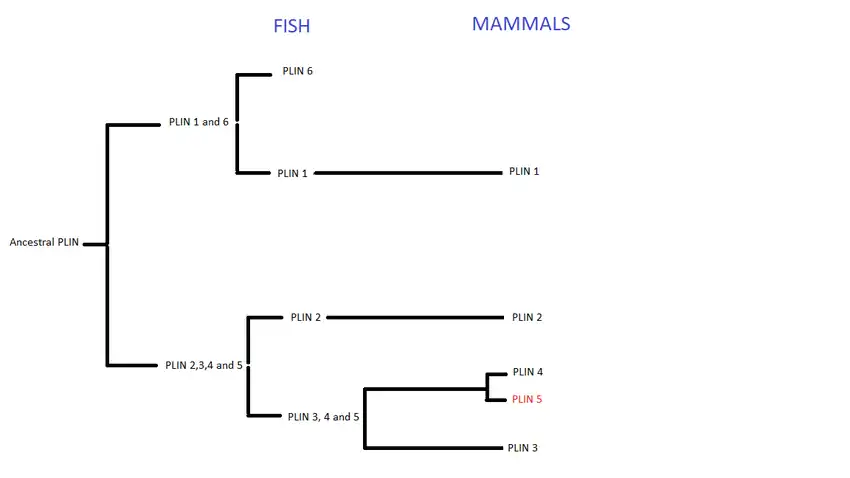
Perilipins are considered to have evolved from a common ancestral gene. This family began to split during the first and second vertebrate genome duplication,[9] giving birth to six types of PLIN genes, expressed throughout the animal kingdom. However, not all types are present in all animals. In fish, PLIN 1 to 6 can be found, whereas in mammals only PLIN1 to 5.[10]
Function
Perilipin 5 is a protein often found in the adipose tissue, especially in those with high oxidative stress, including the heart, liver, skeletal muscle and brown adipose tissue (BAT).[11][12] The perilipin family contributes to the creation of lipid droplets and it also plays a pivotal role in determining what the lipid droplet's function is within the cell.[13] In addition, perilipin 5 regulates the activation of hepatic stellate cell, implicated in fibrosis, which is the creation of new tissue to repair the one damaged.[14]
Beta oxidation
PLIN5, as well as the other members of this protein family, is involved in lipid storage and also has energetic functions. Perilipin 5 coats the lipid droplet acting as a barrier to triglyceride, thus increasing its storage.[15] Under basal condition, PLIN5 decreases lipolysis to prevent energy waste. If energy is needed, perilipin facilitates lipid droplets with lipases and promotes enzymatic activity, regulating energy consumption.[16]
Perilipin 5-Nucleus interactions
The mechanism by which OXPAT5 balances energy remain to be fully clarified. It interacts mainly with LDs, though recent data suggest that this protein also targets the nucleus. When needed, perilipin 5 is phosphorylated by protein kinase A, which allows it to access the nucleus and enter the complex SIRT1/PGC-1α, involved in fatty acid oxidation. PLIN5 removes SIRT1 inhibitor DBC1, therefore increasing its activity. As a consequence, PGC-1α boost activity levels activating certain types of genes that enhances mitochondrial function.[17]
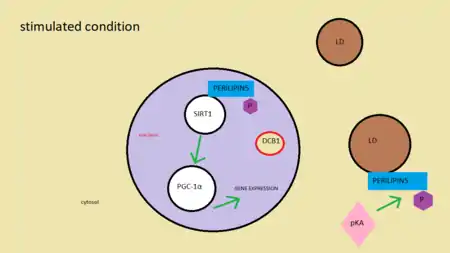
Mitochondria and Perilipin 5
Higher levels of perilipin 5 are closely related to the association between lipid droplet and mitochondria. This is useful in case of starvation (energy shortage). When energy is needed and no glucose is available, the cell uses lipids to feed itself. In order to do that, lipid droplets move towards the mitochondria to transfer the stored fatty acids. The perilipin family opens the channel, giving way to lipids to access the mitochondria. On the other hand, this association may occur with the aim of protecting the mitochondria against toxic levels of fatty acids in the cytosol.[18]
Mean Corpuscular Volume
PLIN5 is an enhancer of the erythrocytes count in blood, as well as hemoglobin concentration.[19]
Lipid Droplet Formation
Mammalian Plin's are not necessary for Lipid Droplet biogenesis, but as they are the primary regulators of lipolysis, they control cellular TAG/CE levels, long-chains whose functions is to provide enough metabolic precursors as polar lipids, which will then create Lipid Droplets.
Clinical importance
PLIN5 is an important regulator of cardiac and liver LDs. Both overexpression and deficiency result in serious consequences. Nevertheless, insights into PLIN5 function may contribute to therapeutic strategies that seek to exploit thermogenic adipose tissue; for example, promoting PLIN5 expression in mouse brown adipose tissue is associated with healthy remodeling of subcutaneous white adipose tissue as well as improvements in systemic glucose tolerance.[20]
Overexpression
Perilipin 5 overexpression causes LD enlargement, accumulation of triglycerides (TAG) and mitochondria dysfunction, causing severe health issues.
Cardiac steatosis
An increase in PLIN5 expression leads to the accumulation of triglyceride content and to the enlargement of LDs and a reduction in its number. This results in cardiac steatosis, an abnormal retention of lipids within a cell. Despite massive steatosis, overexpression of cardiac PLIN5 is compatible with normal heart function and lifespan.[21] Overexpression of PLIN5 also results in concentric hypertrophia on the left ventricle.[22]
Change in mitochondrial morphology
Overexpression of PLIN5 causes a change in mitochondrial function as it leads to the "recruitment" of mitochondria to the LD and changes its morphology. Such LD-mitochondria assemblies typically move in unison due to their tight association. This mitochondrial phenotype was only observed in close proximity to LDs.[23][24]
Fatty liver disease
PLIN5 plays an important role in regulating lipid accumulation and its breakdown in the liver. It appears that statins, a class of lipid-lowering medication, were effective in treating fatty liver diseases in non-alcoholic patients, as it decreased the hepatic expression of PLIN5, thus decreasing lipid accumulation.[25]
Deficiency
When LDs are lost due to a deficiency in PLIN5, FAs are not sequestered as TAG in LD and therefore larger amounts of FAs are oxidized in the mitochondria leading to an excess generation of reactive oxygen species (ROS). Medium and high concentrations of ROS can induce apoptosis and eventually cause necrosis through oxidative stress.[26] PLIN deficiency also reduces superoxide dismutase (SOD) activity. Furthermore, deficiency in PLIN5 initiates excessive phosforilation of PI3K/Akt which contributes to ischemia-reperfusion injury aggravation.
PLIN5-knockout diabetic mice might avoid excess accumulation of lipotoxic molecules such as DAG and ceramide, a common problem among diabetic patients. The built up of DAG and ceramide disrupts several signaling pathways, including PKC pathway. However, in humans, the association is not clear and the packaging of TAG into lipid droplets may be of critical importance in determining whether the accumulation of excess lipids exert a toxic effect on the myocardium.
It was found that a deficiency in Plin5 reduces mitochondrial oxidative capacity in mouses. Mitochondria from hearts suffering from a Plin5 deficiency had a membrana whose fatty acyl composition was altered, and its depolarization was compromised.[27]
It was also discovered in mice that, if a whole body deficiency of Plin5 was to happen, cardiac lipid droplet formation ability would be reduced, increasing fatty acid oxidations and promoting cardiac dysfunction. This, however, could be prevented by anti-oxidative therapy.
Atherosclerosis
Deficiency in PLIN5 increases monocytes which are critical participants in the inflammation process. Because inflammation is essential to atherosclerosis, a deficiency in PLIN5 increases atherosclerosis risk.[28]
See also
External links
References
- 1 2 3 GRCh38: Ensembl release 89: ENSG00000214456 - Ensembl, May 2017
- 1 2 3 GRCm38: Ensembl release 89: ENSMUSG00000011305 - Ensembl, May 2017
- ↑ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ↑ "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ↑ Lipid droplet biogenesis, structure and degradation are regulated, Hashani M, Witzel HR, Pawella LM, Lehmann-Koch J
- ↑ Perilipin 5, a lipid droplet protein adapted to mitochondrial energy utilization. Kimmel AR, Sztalryd C.
- ↑ R. Mason, Rachael (2014). "PLIN5 deletion remodels intracellular lipid composition and causes insulin resistance in muscle". Molecular Metabolism. 3 (6): 652–63. doi:10.1016/j.molmet.2014.06.002. PMC 4142393. PMID 25161888.
- ↑ Widespread expression of perilipin 5 in normal human tissues and in diseases is restricted to distinct lipid droplet subpopulations. Hashani M, Witzel HR, Pawella LM
- ↑ Dehal, Paramvir (2005-09-06). "Two Rounds of Whole Genome Duplication in the Ancestral Vertebrate". PLOS Biology. 3 (10): e314. doi:10.1371/journal.pbio.0030314. PMC 1197285. PMID 16128622.
- ↑ Granneman, James G. (2017-02-28). "Lipid droplet biology and evolution illuminated by the characterization of a novel perilipin in teleost fish". eLife. 6. doi:10.7554/eLife.21771. PMC 5342826. PMID 28244868.
- ↑ Alan R., Kimmel (2014-04-25). "Perilipin 5, a Lipid Droplet Protein Adapted to Mitochondrial Energy Utilization". Current Opinion in Lipidology. 25 (2): 110–7. doi:10.1097/MOL.0000000000000057. PMC 4517968. PMID 24535284.
- ↑ PL, Zhou (2017-12-07). "Plin5 deficiency promotes atherosclerosis progression through accelerating inflammation, apoptosis and oxidative stress". Journal of Cellular Biochemistry. doi:10.1002/jcb.26493. PMID 29215758. S2CID 36675962.
- ↑ Kenta, Kuramoto (2012-07-06). "Perilipin 5, a Lipid Droplet-binding Protein, Protects Heart from Oxidative Burden by Sequestering Fatty Acid from Excessive Oxidation". Journal of Biological Chemistry. 287 (28): 23852–63. doi:10.1074/jbc.M111.328708. PMC 3390660. PMID 22532565.
- ↑ Jianguo, Lin (2016-05-02). "Perilipin 5 restores the formation of lipid droplets in activated hepatic stellate cells and inhibits their activation". Laboratory Investigation. 96 (7): 791–806. doi:10.1038/labinvest.2016.53. PMID 27135793.
- ↑ Andersson, Linda (2017-07-21). "Deficiency in perilipin 5 reduces mitochondrial function and membrane depolarization in mouse hearts". The International Journal of Biochemistry & Cell Biology. 91 (Pt A): 9–13. doi:10.1016/j.biocel.2017.07.021. PMID 28811250.
- ↑ Dawn L., Brasaemle (2007-09-18). "Thematic review series: adipocyte biology. The perilipin family of structural lipid droplet proteins: stabilization of lipid droplets and control of lipolysis". Journal of Lipid Research. 48 (12): 2547–59. doi:10.1194/jlr.R700014-JLR200. PMID 17878492.
- ↑ Gallardo-Montejano, Violeta I. (2016-08-24). "Nuclear Perilipin 5 integrates lipid droplet lipolysis with PGC-1α/SIRT1-dependent transcriptional regulation of mitochondrial function". Nature Communications. 7: 12723. Bibcode:2016NatCo...712723G. doi:10.1038/ncomms12723. PMC 4999519. PMID 27554864.
- ↑ Alan R., Kimmel (2014-04-25). "Perilipin 5, a Lipid Droplet Protein Adapted to Mitochondrial Energy Utilization". Current Opinion in Lipidology. 25 (2): 110–7. doi:10.1097/MOL.0000000000000057. PMC 4517968. PMID 24535284.
- ↑ Astle, William J.; Elding, Heather; Jiang, Tao; Allen, Dave; Ruklisa, Dace; Mann, Alice L.; Mead, Daniel; Bouman, Heleen; Riveros-Mckay, Fernando (2016-11-17). "The Allelic Landscape of Human Blood Cell Trait Variation and Links to Common Complex Disease". Cell. 167 (5): 1415–1429.e19. doi:10.1016/j.cell.2016.10.042. ISSN 1097-4172. PMC 5300907. PMID 27863252.
- ↑ Gallardo-Montejano, Violeta I.; Yang, Chaofeng; Hahner, Lisa; McAfee, John L.; Johnson, Joshua A.; Holland, William L.; Fernandez-Valdivia, Rodrigo; Bickel, Perry E. (2021). "Perilipin 5 links mitochondrial uncoupled respiration in brown fat to healthy white fat remodeling and systemic glucose tolerance". Nature Communications. 12 (1): 3320. Bibcode:2021NatCo..12.3320G. doi:10.1038/s41467-021-23601-2. ISSN 2041-1723. PMC 8175597. PMID 34083525.
- ↑ Pollak, Nina M. (2015-01-16). "The Interplay of Protein Kinase A and Perilipin 5 Regulates Cardiac Lipolysis". Journal of Biological Chemistry. 290 (3): 1295–306. doi:10.1074/jbc.M114.604744. PMC 4340377. PMID 25418045.
- ↑ Wang, Hong (2013). "Cardiomyocyte-specific perilipin 5 overexpression leads to myocardial steatosis and modest cardiac dysfunction". Journal of Lipid Research. 54 (4): 953–65. doi:10.1194/jlr.M032466. PMC 3606001. PMID 23345411.
- ↑ Pribasnig, Maria (2018-07-08). "Extended-resolution imaging of the interaction of lipid droplets and mitochondria". Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids. 1863 (10): 1285–1296. doi:10.1016/j.bbalip.2018.07.008. PMID 30305245.
- ↑ Benador, IY; Veliova, M; Mahdaviani, K; Petcherski, A; Wikstrom, JD; Assali, EA; Acín-Pérez, R; Shum, M; Oliveira, MF; Cinti, S; Sztalryd, C; Barshop, WD; Wohlschlegel, JA; Corkey, BE; Liesa, M; Shirihai, OS (3 April 2018). "Mitochondria Bound to Lipid Droplets Have Unique Bioenergetics, Composition, and Dynamics that Support Lipid Droplet Expansion". Cell Metabolism. 27 (4): 869–885.e6. doi:10.1016/j.cmet.2018.03.003. PMC 5969538. PMID 29617645.
- ↑ Cédric, Langhi (2014-04-09). "Perilipin-5 is regulated by statins and controls triglyceride contents in the hepatocyte". Journal of Hepatology. 61 (2): 358–65. doi:10.1016/j.jhep.2014.04.009. PMC 4104237. PMID 24768901.
- ↑ Zheng, Pengfei (2017-02-20). "Plin5 alleviates myocardial ischaemia/reperfusion injury by reducing oxidative stress through inhibiting the lipolysis of lipid droplets". Scientific Reports. 7: 42574. Bibcode:2017NatSR...742574Z. doi:10.1038/srep42574. PMC 5316932. PMID 28218306.
- ↑ Andersson, Linda; Drevinge, Christina; Mardani, Ismena; Dalen, Knut T.; Ståhlman, Marcus; Klevstig, Martina; Lundqvist, Annika; Haugen, Fred; Adiels, Martin (2017-08-12). "Deficiency in perilipin 5 reduces mitochondrial function and membrane depolarization in mouse hearts". The International Journal of Biochemistry & Cell Biology. 91 (Pt A): 9–13. doi:10.1016/j.biocel.2017.07.021. ISSN 1878-5875. PMID 28811250.
- ↑ Zhou, PL (2017-12-07). "Plin5 deficiency promotes atherosclerosis progression through accelerating inflammation, apoptosis and oxidative stress". Journal of Cellular Biochemistry. doi:10.1002/jcb.26493. PMID 29215758. S2CID 36675962.