| Siboglinidae Temporal range: | |
|---|---|
 | |
| Riftia pachyptila | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Annelida |
| Clade: | Pleistoannelida |
| Subclass: | Sedentaria |
| Infraclass: | Canalipalpata |
| Order: | Sabellida |
| Family: | Siboglinidae Caullery, 1914 |
| Genera | |
Siboglinidae is a family of polychaete annelid worms whose members made up the former phyla Pogonophora and Vestimentifera (the giant tube worms).[1][2] The family is composed of around 100 species of vermiform creatures which live in thin tubes buried in sediment (Pogonophora) or in tubes attached to hard substratum (Vestimentifera) at ocean depths ranging from 100 to 10,000 m (300 to 32,800 ft). They can also be found in association with hydrothermal vents, methane seeps, sunken plant material, and whale carcasses.
The first specimen was dredged from the waters of Indonesia in 1900. These specimens were given to French zoologist Maurice Caullery, who studied them for nearly 50 years.
Anatomy
Most siboglinids are less than 1 millimetre (0.04 in) in diameter, but 10–75 centimetres (3.9–29.5 in) in length. They inhabit tubular structures composed of chitin which are fixed to rocks or substrates. The tubes are often clustered together in large colonies.[3]
Their bodies are divided into four regions. The anterior end is called the cephalic lobe, which ranges from one to over 200 thin branchial ciliated tentacles, each with tiny side branches known as pinnules. Behind this is a glandular forepart, which helps to secrete the tube. The main part of the body is the trunk, which is greatly elongated and bears various annuli, papillae, and ciliary tracts. Posterior to the trunk is the short metamerically segmented opisthosoma, bearing external paired chaetae, which help to anchor the animal to the base of its tube.[3]
The body cavity has a separate compartment in each of the first three regions of the body and extends into the tentacles. The opisthosoma has a coelomic chamber in each of its 5 to 23 segments, separated by septa. The worms have a complex closed circulatory system and a well-developed nervous system, but as adults, siboglinids completely lack a mouth, gut, and anus.[4]
Evolution
The family Siboglinidae has been difficult to place in an evolutionary context.[5] After examination of genetic differences between annelids, Siboglinidae were placed within the order Polychaeta by scientific consensus.[6] The fossil record along with molecular clocks suggest the family has Mesozoic (250 – 66 Mya) or Cenozoic (66 Mya – recent) origins.[5] However, some fossils of crystallized tubes are attributed to early Siboglinidae dating back to 500 Mya.[5] Molecular work aligning five genes has identified four distinct clades within Siboglinidae.[7][8][9] The clades are Vestimentifera, Sclerolinum, Frenulata, and Osedax.[8] Vestimentiferans live in vent and seep habitats.[8] Separation of vestimentiferans into seep and deep-sea-dwelling clades is still debated due to some phylogenies based on sequencing data placing the genera along a continuum.[10] Sclerolinum is a monogeneric clade (which may be called Monilifera) living on organic-rich remains.[5] Frenulates live in organic-rich sediment habitats.[11] Osedax is a monogeneric clade specialized in living on whale bones, although recent evidence shows them living on fish bones as well.[12]
One probable relationship between the four clades is shown in the cladogram below. The position of Osedax is weakly supported.[5]
| Siboglinidae |
| ||||||||||||||||||
Vestimentiferans
Like other tube worms, vestimentiferans are benthic marine creatures. Riftia pachyptila, a vestimentiferan, is known only from the hydrothermal vent systems.[5]
Anatomy of vestimentiferans
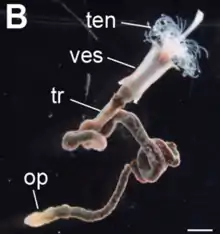
Vestimentiferan bodies are divided into four regions: the obturaculum, vestimentum, trunk, and opisthosome. The main trunk of the body bears wing-like extensions. Unlike other siboglinids that never have a digestive tract, they have one that they completely lose during metamorphosis.
The obturaculum is the first anterior body part.[13] It is possible that the obturaculum is actually an outgrowth of the vestimentum rather than a separate body segment which would distinguish it from other siboglinids.
The vestimentum, from which the group's name is derived, is a wing-like body part with glands that secrete the tube. In a ventroanterior position in the vestimentum is the brain which is postulated to be simpler than relatives that maintain a gut in the adult form.[13] The opisthosome is the anchoring rear body part.
Vestimentiferan ecology
Their primary nutrition is derived from the sulfide-rich fluids emanating from the hydrothermal vents where they live. The sulfides are metabolized by symbiotic hydrogen sulfide- or methane-oxidizing bacteria living in an internal organ, the trophosome. One gram of trophosome tissue can contain one billion bacteria. The origin of this symbiotic relationship is not currently known. The bacteria appear to colonize the host animal larvae after they have settled on a surface, entering them through their skin.[14] This method of entry, known as horizontal transmission, means that each organism may have different species of bacteria assisting in this symbiosis. However, these bacteria all play similar roles in sustaining the vestimentiferans. Endosymbionts have a wide variety of metabolic genes, which may allow them to switch between autotrophic and heterotrophic methods of nutrient acquisition.[15] When the host dies, the bacteria are released and return to the free-living population in the seawater.[16]
Discovery of the hydrothermal vents in the eastern Pacific Ocean was quickly followed by the discovery and description of new vestimentiferan tubeworm species. These tubeworms are one of the most dominant organisms associated with the hydrothermal vents in the Pacific Ocean. Tubeworms anchor themselves to the substratum of the hydrocarbon seep by roots located at the basal portion of their bodies.[17] Intact tubeworm roots have proven very difficult to obtain for study because they are extremely delicate, and often break off when a tubeworm is removed from hypothermal vent regions. How long the roots of the tube worms can grow is unknown, but roots have been recovered longer than 30 m.
A single aggregation of tubeworms can contain thousands of individuals, and the roots produced by each tubeworm can become tangled with the roots of neighbouring tubeworms.[18] These mats of roots are known as "ropes", and travel down the tubes of dead tubeworms, and run through holes in rocks. The diameter and wall thickness of the tubeworm roots do not appear to change with distance from the trunk portion of the tubeworm's body.
Like the trunk portion of the body, the roots of the vestimentiferan tubeworms are composed of chitin crystallites, which support and protect the tubeworm from predation and environmental stresses. Tubeworms build the external chitin structure themselves by secreting chitin from specialized glands located in their body walls.
Genera
- Osedax
- Clade Frenulata[2]
- Birsteinia
- Bobmarleya[19]
- Choanophorus
- Crassibrachia
- Cyclobrachia
- Diplobrachia
- Galathealinum
- Heptabrachia
- Lamellisabella
- Nereilinum
- Oligobrachia
- Paraescarpia[20]
- Polybrachia
- Siboglinoides
- Siboglinum
- Siphonobrachia
- Spirobrachia
- Unibrachium
- Volvobrachia[21]
- Zenkevitchiana
- Clade Monilifera[22]
- Sclerolinum[2]
- Clade Vestimentifera[2]
- Alaysia
- Arcovesia
- Escarpia
- Lamellibrachia
- Oasisia
- Ridgeia
- Riftia
- Tevnia
References
- ↑ Kojima, S.; Hashimoto, T.; Hasegawa, M.; Murata, S.; Ohta, S.; Seki, H.; Okada, N. (July 1993). "Close phylogenetic relationship between vestimentifera (tube worms) and annelida revealed by the amino acid sequence of elongation factor-lα". Journal of Molecular Evolution. 37 (1): 66–70. Bibcode:1993JMolE..37...66K. doi:10.1007/BF00170463. PMID 8360920. S2CID 23391565.
- 1 2 3 4 Rouse, G. W. (2001). "A cladistic analysis of Siboglinidae Caullery, 1914 (Polychaeta, Annelida): formerly the phyla Pogonophora and Vestimentifera". Zoological Journal of the Linnean Society. 132 (1): 55–80. doi:10.1006/zjls.2000.0263.
- 1 2 Barnes, Robert D. (1982). Invertebrate Zoology. Philadelphia, PA: Holt-Saunders International. pp. 862–863. ISBN 0-03-056747-5.
- ↑ Ross Piper (30 August 2007). Extraordinary animals: an encyclopedia of curious and unusual animals. Greenwood Publishing Group. ISBN 978-0-313-33922-6. Retrieved 3 December 2011.
- 1 2 3 4 5 6 Hilário, Ana; Capa, María; Dahlgren, Thomas G.; Halanych, Kenneth M.; Little, Crispin T. S.; Thornhill, Daniel J.; Verna, Caroline; Glover, Adrian G. (2011). Laudet, Vincent (ed.). "New Perspectives on the Ecology and Evolution of Siboglinid Tubeworms". PLOS ONE. 6 (2): e16309. Bibcode:2011PLoSO...616309H. doi:10.1371/journal.pone.0016309. PMC 3038861. PMID 21339826.
- ↑ Struck, T. H.; Schult, N.; Kusen, T.; Hickman, E.; Bleidorn, C.; McHugh, D.; Halanych, K. M. (2007). "Annelid phylogeny and the status of Sipuncula and Echiura". BMC Evolutionary Biology. 7: 57. doi:10.1186/1471-2148-7-57. PMC 1855331. PMID 17411434.
- ↑ Black, M. B.; Halanych, K. M.; Maas, P. A. Y.; Hoeh, W. R.; Hashimoto, J.; Desbruyeres, D.; Lutz, R. A.; et al. (1997). "Molecular systematics of vestimentiferan tubeworms from hydrothermal vents and cold-water seeps". Marine Biology. 130 (2): 141–149. doi:10.1007/s002270050233. S2CID 13217485.
- 1 2 3 Glover, A. G.; Kallstrom, B.; Smith, C. R.; Dahlgren, T. G. (2005). "World-wide whale worms? A new species of Osedax from the shallow north Atlantic". Proceedings of the Royal Society B: Biological Sciences. 272 (1581): 2587–2592. doi:10.1098/rspb.2005.3275. PMC 1559975. PMID 16321780.
- ↑ Vrijenhoek, R. C.; Johnson, S. B.; Rouse, G. W. (2009). "A remarkable diversity of bone-eating worms (Osedax; Siboglinidae; Annelida)". BMC Biology. 7: 74. doi:10.1186/1741-7007-7-74. PMC 2780999. PMID 19903327.
- ↑ Bright, Monika; Lallie, François (2010-05-12), Gibson, R; Atkinson, R; Gordon, J (eds.), "The Biology of Vestimentiferan Tubeworms", Oceanography and Marine Biology, CRC Press, vol. 20103650, pp. 213–265, doi:10.1201/ebk1439821169-c4, ISBN 9781439821169
- ↑ Rodrigues, C. F.; Hilário, A.; Cunha, M. R.; Weightman, A. J.; Webster, G. (2011). "Microbial diversity in Frenulata (Siboglinidae, Polychaeta) species from mud volcanoes in the Gulf of Cadiz (NE Atlantic)". Antonie van Leeuwenhoek. 100 (1): 83–98. doi:10.1007/s10482-011-9567-0. PMID 21359663. S2CID 10224623.
- ↑ Rouse, G. W.; Goffredi, S. K.; Johnson, S. B.; Vrijenhoek, R. C. (2011). "Not whale-fall specialists, Osedax worms also consume fishbones". Biology Letters. 7 (5): 736–739. doi:10.1098/rsbl.2011.0202. PMC 3169056. PMID 21490008.
- 1 2 Miyamoto, Norio; Shinozaki, Ayuta; Fujiwara, Yoshihiro (2013-01-23). "Neuroanatomy of the Vestimentiferan Tubeworm Lamellibrachia satsuma Provides Insights into the Evolution of the Polychaete Nervous System". PLOS ONE. 8 (1): e55151. doi:10.1371/journal.pone.0055151. PMC 3553155. PMID 23372830.
- ↑ Nussbaumer, Andrea D.; Fisher, Charles R.; Bright, Monika (2006-05-18). "Horizontal endosymbiont transmission in hydrothermal vent tubeworms". Nature. 441 (7091): 345–348. Bibcode:2006Natur.441..345N. doi:10.1038/nature04793. ISSN 1476-4687. PMID 16710420. S2CID 18356960.
- ↑ Reveillaud, Julie; Anderson, Rika; Reves-Sohn, Sintra; Cavanaugh, Colleen; Huber, Julie A. (2018-01-27). "Metagenomic investigation of vestimentiferan tubeworm endosymbionts from Mid-Cayman Rise reveals new insights into metabolism and diversity". Microbiome. 6 (1): 19. doi:10.1186/s40168-018-0411-x. ISSN 2049-2618. PMC 5787263. PMID 29374496.
- ↑ Klose, Julia; Polz, Martin F.; Wagner, Michael; Schimak, Mario P.; Gollner, Sabine; Bright, Monika (2015-09-08). "Endosymbionts escape dead hydrothermal vent tubeworms to enrich the free-living population". Proceedings of the National Academy of Sciences of the United States of America. 112 (36): 11300–11305. Bibcode:2015PNAS..11211300K. doi:10.1073/pnas.1501160112. ISSN 1091-6490. PMC 4568656. PMID 26283348.
- ↑ Halanych, K.; Maas, P.; Hoeh, W.; Hashimoto, J.; Desbruyeres, D.; Lutz, R.; Vrijenhoek, R. (1997). "Molecular systematics of vestimentiferan tubeworms from hydrothermal vents and cold-water seeps". Marine Biology. 130 (2): 141–149. doi:10.1007/s002270050233. S2CID 13217485.
- ↑ Julian, D.; Gaill, F.; Wood, E.; Arp, A.; Fisher, C. (1999). "Roots as a site of hydrogen sulphide uptake in the hydrocarbon seep vestimentiferan Lamellibrachia sp". The Journal of Experimental Biology. 202 (Pt 17): 2245–57. doi:10.1242/jeb.202.17.2245. PMID 10441078.
- ↑ Hilário, A.; Cunha, M. R. (2008). "On some frenulate species (Annelida: Polychaeta: Siboglinidae) from mud volcanoes in the Gulf of Cadiz (NE Atlantic)". Scientia Marina. 72 (2): 361–371. doi:10.3989/scimar.2008.72n2361.
- ↑ Southward, E. C.; Schulze, A.; Tunnicliffe, V. (2002). "Vestimentiferans (Pogonophora) in the Pacific and Indian Oceans: a new genus from Lihir Island (Papua New Guinea) and the Java Trench, with the first report of Arcovestia ivanovi from the North Fiji Basin". Journal of Natural History. 36 (10): 1179–1197. doi:10.1080/00222930110040402. S2CID 86076917.
- ↑ Smirnov, R. V. (2000). "A redescription of Spirobrachia leospira Gureeva (Pogonophora), with the erection of a new genus and a revision of the Spirobrachiidae". Ophelia. 53 (2): 151–158. doi:10.1080/00785236.2000.10409445. S2CID 85319259.
- ↑ Halanych, Kenneth M. (2005). "Molecular phylogeny of siboglinid annelids (a.k.a. pogonophorans): a review". Hydrobiologia. 535–536 (1): 297–307. doi:10.1007/s10750-004-1437-6. ISSN 0018-8158. S2CID 16022909.
External links
 Media related to Siboglinidae at Wikimedia Commons
Media related to Siboglinidae at Wikimedia Commons Data related to Siboglinidae at Wikispecies
Data related to Siboglinidae at Wikispecies