| rRNAs | |
|---|---|
 rRNAs of various species | |
| Identifiers | |
| Other data | |
| RNA type | Gene; rRNA |
| PDB structures | PDBe |
Ribosomal ribonucleic acid (rRNA) is a type of non-coding RNA which is the primary component of ribosomes, essential to all cells. rRNA is a ribozyme which carries out protein synthesis in ribosomes. Ribosomal RNA is transcribed from ribosomal DNA (rDNA) and then bound to ribosomal proteins to form small and large ribosome subunits. rRNA is the physical and mechanical factor of the ribosome that forces transfer RNA (tRNA) and messenger RNA (mRNA) to process and translate the latter into proteins.[1] Ribosomal RNA is the predominant form of RNA found in most cells; it makes up about 80% of cellular RNA despite never being translated into proteins itself. Ribosomes are composed of approximately 60% rRNA and 40% ribosomal proteins by mass.
Structure
Although the primary structure of rRNA sequences can vary across organisms, base-pairing within these sequences commonly forms stem-loop configurations. The length and position of these rRNA stem-loops allow them to create three-dimensional rRNA structures that are similar across species.[2] Because of these configurations, rRNA can form tight and specific interactions with ribosomal proteins to form ribosomal subunits. These ribosomal proteins contain basic residues (as opposed to acidic residues) and aromatic residues (i.e. phenylalanine, tyrosine and tryptophan) allowing them to form chemical interactions with their associated RNA regions, such as stacking interactions. Ribosomal proteins can also cross-link to the sugar-phosphate backbone of rRNA with binding sites that consist of basic residues (i.e. lysine and arginine). All ribosomal proteins (including the specific sequences that bind to rRNA) have been identified. These interactions along with the association of the small and large ribosomal subunits result in a functioning ribosome capable of synthesizing proteins.[3]
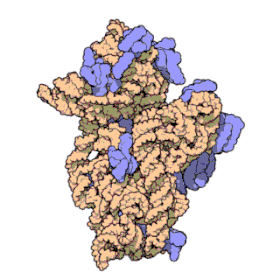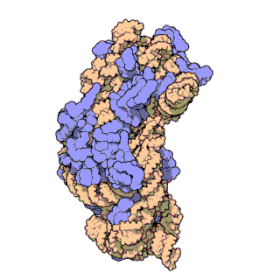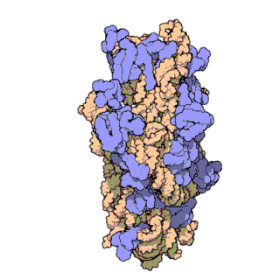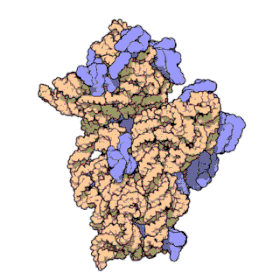
Ribosomal RNA organizes into two types of major ribosomal subunit: the large subunit (LSU) and the small subunit (SSU). One of each type come together to form a functioning ribosome. The subunits are at times referred to by their size-sedimentation measurements (a number with an "S" suffix). In prokaryotes, the LSU and SSU are called the 50S and 30S subunits, respectively. In eukaryotes, they are a little larger; the LSU and SSU of eukaryotes are termed the 60S and 40S subunits, respectively.
In the ribosomes of prokaryotes such as bacteria, the SSU contains a single small rRNA molecule (~1500 nucleotides) while the LSU contains one single small rRNA and a single large rRNA molecule (~3000 nucleotides). These are combined with ~50 ribosomal proteins to form ribosomal subunits. There are three types of rRNA found in prokaryotic ribosomes: 23S and 5S rRNA in the LSU and 16S rRNA in the SSU.
In the ribosomes of eukaryotes such as humans, the SSU contains a single small rRNA (~1800 nucleotides) while the LSU contains two small rRNAs and one molecule of large rRNA (~5000 nucleotides). Eukaryotic rRNA has over 70 ribosomal proteins which interact to form larger and more polymorphic ribosomal units in comparison to prokaryotes.[4] There are four types of rRNA in eukaryotes: 3 species in the LSU and 1 in the SSU.[5] Yeast has been the traditional model for observation of eukaryotic rRNA behavior and processes, leading to a deficit in diversification of research. It has only been within the last decade that technical advances (specifically in the field of Cryo-EM) have allowed for preliminary investigation into ribosomal behavior in other eukaryotes.[6] In yeast, the LSU contains the 5S, 5.8S and 28S rRNAs. The combined 5.8S and 28S are roughly equivalent in size and function to the prokaryotic 23S rRNA subtype, minus expansion segments (ESs) that are localized to the surface of the ribosome which were thought to occur only in eukaryotes. However recently, the Asgard phyla, namely, Lokiarchaeota and Heimdallarchaeota, considered the closest archaeal relatives to Eukarya, were reported to possess two supersized ESs in their 23S rRNAs.[7] Likewise, the 5S rRNA contains a 108‐nucleotide insertion in the ribosomes of the halophilic archaeon Halococcus morrhuae.[8][9]
A eukaryotic SSU contains the 18S rRNA subunit, which also contains ESs. SSU ESs are generally smaller than LSU ESs.
SSU and LSU rRNA sequences are widely used for study of evolutionary relationships among organisms, since they are of ancient origin,[10] are found in all known forms of life and are resistant to horizontal gene transfer. rRNA sequences are conserved (unchanged) over time due to their crucial role in the function of the ribosome.[11] Phylogenic information derived from the 16s rRNA is currently used as the main method of delineation between similar prokaryotic species by calculating nucleotide similarity.[12] The canonical tree of life is the lineage of the translation system.
LSU rRNA subtypes have been called ribozymes because ribosomal proteins cannot bind to the catalytic site of the ribosome in this area (specifically the peptidyl transferase center, or PTC).[13]
The SSU rRNA subtypes decode mRNA in its decoding center (DC).[14] Ribosomal proteins cannot enter the DC.
The structure of rRNA is able to drastically change to affect tRNA binding to the ribosome during translation of other mRNAs.[15] In 16S rRNA, this is thought to occur when certain nucleotides in the rRNA appear to alternate base pairing between one nucleotide or another, forming a "switch" that alters the rRNA's conformation. This process is able to affect the structure of the LSU and SSU, suggesting that this conformational switch in the rRNA structure affects the entire ribosome in its ability to match a codon with its anticodon in tRNA selection as well as decode mRNA.[16]
Assembly
Ribosomal RNA's integration and assembly into ribosomes begins with their folding, modification, processing and assembly with ribosomal proteins to form the two ribosomal subunits, the LSU and the SSU. In Prokaryotes, rRNA incorporation occurs in the cytoplasm due to the lack of membrane-bound organelles. In Eukaryotes, however, this process primarily takes place in the nucleolus and is initiated by the synthesis of pre-RNA. This requires the presence of all three RNA polymerases. In fact, the transcription of pre-RNA by RNA polymerase I accounts for about 60% of cell's total cellular RNA transcription.[17] This is followed by the folding of the pre-RNA so that it can be assembled with ribosomal proteins. This folding is catalyzed by endo- and exonucleases, RNA helicases, GTPases and ATPases. The rRNA subsequently undergoes endo- and exonucleolytic processing to remove external and internal transcribed spacers.[18] The pre-RNA then undergoes modifications such as methylation or pseudouridinylation before ribosome assembly factors and ribosomal proteins assemble with the pre-RNA to form pre-ribosomal particles. Upon going under more maturation steps and subsequent exit from the nucleolus into the cytoplasm, these particles combine to form the ribosomes.[18] The basic and aromatic residues found within the primary structure of rRNA allow for favorable stacking interactions and attraction to ribosomal proteins, creating a cross-linking effect between the backbone of rRNA and other components of the ribosomal unit. More detail on the initiation and beginning portion of these processes can be found in the "Biosynthesis" section.
Function
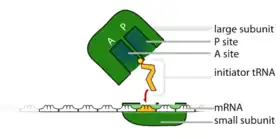
Universally conserved secondary structural elements in rRNA among different species show that these sequences are some of the oldest discovered. They serve critical roles in forming the catalytic sites of translation of mRNA. During translation of mRNA, rRNA functions to bind both mRNA and tRNA to facilitate the process of translating mRNA's codon sequence into amino acids. rRNA initiates the catalysis of protein synthesis when tRNA is sandwiched between the SSU and LSU. In the SSU, the mRNA interacts with the anticodons of the tRNA. In the LSU, the amino acid acceptor stem of the tRNA interacts with the LSU rRNA. The ribosome catalyzes ester-amide exchange, transferring the C-terminus of a nascent peptide from a tRNA to the amine of an amino acid. These processes are able to occur due to sites within the ribosome in which these molecules can bind, formed by the rRNA stem-loops. A ribosome has three of these binding sites called the A, P and E sites:
- In general, the A (aminoacyl) site contains an aminoacyl-tRNA (a tRNA esterified to an amino acid on the 3' end).
- The P (peptidyl) site contains a tRNA esterified to the nascent peptide. The free amino (NH2) group of the A site tRNA attacks the ester linkage of P site tRNA, causing transfer of the nascent peptide to the amino acid in the A site. This reaction is takes place in the peptidyl transferase center[13]
- The E (exit) site contains a tRNA that has been discharged, with a free 3' end (with no amino acid or nascent peptide).
A single mRNA can be translated simultaneously by multiple ribosomes. This is called a polysome.
In prokaryotes, much work has been done to further identify the importance of rRNA in translation of mRNA. For example, it has been found that the A site consists primarily of 16S rRNA. Apart from various protein elements that interact with tRNA at this site, it is hypothesized that if these proteins were removed without altering ribosomal structure, the site would continue to function normally. In the P site, through the observation of crystal structures it has been shown the 3' end of 16s rRNA can fold into the site as if a molecule of mRNA. This results in intermolecular interactions that stabilize the subunits. Similarly, like the A site, the P site primarily contains rRNA with few proteins. The peptidyl transferase center, for example, is formed by nucleotides from the 23S rRNA subunit.[13] In fact, studies have shown that the peptidyl transferase center contains no proteins, and is entirely initiated by the presence of rRNA. Unlike the A and P sites, the E site contains more proteins. Because proteins are not essential for the functioning of the A and P sites, the E site molecular composition shows that it is perhaps evolved later. In primitive ribosomes, it is likely that tRNAs exited from the P site. Additionally, it has been shown that E-site tRNA bind with both the 16S and 23S rRNA subunits.[19]
Subunits and associated ribosomal RNA

Both prokaryotic and eukaryotic ribosomes can be broken down into two subunits, one large and one small. The exemplary species used in the table below for their respective rRNAs are the bacterium Escherichia coli (prokaryote) and human (eukaryote). Note that "nt" represents the length of the rRNA type in nucleotides and the "S" (such as in "16S) represents Svedberg units.
| Type | Size | Large subunit (LSU rRNA) | Small subunit (SSU rRNA) |
| prokaryotic | 70S | 50S (5S : 120 nt, 23S : 2906 nt) | 30S (16S : 1542 nt) |
| eukaryotic (nuclear) | 80S | 60S (5S : 121 nt,[20] 5.8S : 156 nt,[21] 28S : 5070 nt[22]) | 40S (18S : 1869 nt[23]) |
| eukaryotic (mitochondrial) | 55S | 39S 16S (Mitochondrially encoded 16S rRNA : approx. 1,571 nt) | 28S 12S (Mitochondrially encoded 12S rRNA : approx. 955 nt)[24] |
S units of the subunits (or the rRNAs) cannot simply be added because they represent measures of sedimentation rate rather than of mass. The sedimentation rate of each subunit is affected by its shape, as well as by its mass. The nt units can be added as these represent the integer number of units in the linear rRNA polymers (for example, the total length of the human rRNA = 7216 nt).
Gene clusters coding for rRNA are commonly called "ribosomal DNA" or rDNA (note that the term seems to imply that ribosomes contain DNA, which is not the case).
In prokaryotes
In prokaryotes a small 30S ribosomal subunit contains the 16S ribosomal RNA. The large 50S ribosomal subunit contains two rRNA species (the 5S and 23S ribosomal RNAs). Therefore it can be deduced that in both bacteria and archaea there is one rRNA gene that codes for all three rRNA types :16S, 23S and 5S.[25]
Bacterial 16S ribosomal RNA, 23S ribosomal RNA, and 5S rRNA genes are typically organized as a co-transcribed operon. As shown by the image in this section, there is an internal transcribed spacer between 16S and 23S rRNA genes.[26] There may be one or more copies of the operon dispersed in the genome (for example, Escherichia coli has seven). Typically in bacteria there are between one and fifteen copies.[25]
Archaea contains either a single rRNA gene operon or up to four copies of the same operon.[25]
The 3' end of the 16S ribosomal RNA (in a ribosome) recognizes a sequence on the 5' end of mRNA called the Shine-Dalgarno sequence.
In eukaryotes

In contrast, eukaryotes generally have many copies of the rRNA genes organized in tandem repeats. In humans, approximately 300–400 repeats are present in five clusters, located on chromosomes 13 (RNR1), 14 (RNR2), 15 (RNR3), 21 (RNR4) and 22 (RNR5). Diploid humans have 10 clusters of genomic rDNA which in total make up less than 0.5% of the human genome.[27]
It was previously accepted that repeat rDNA sequences were identical and served as redundancies or failsafes to account for natural replication errors and point mutations. However, sequence variation in rDNA (and subsequently rRNA) in humans across multiple chromosomes has been observed, both within and between human individuals. Many of these variations are palindromic sequences and potential errors due to replication.[28] Certain variants are also expressed in a tissue-specific manner in mice.[29]
Mammalian cells have 2 mitochondrial (12S and 16S) rRNA molecules and 4 types of cytoplasmic rRNA (the 28S, 5.8S, 18S, and 5S subunits). The 28S, 5.8S, and 18S rRNAs are encoded by a single transcription unit (45S) separated by 2 internally transcribed spacers. The first spacer corresponds to the one found in bacteria and archaea, and the other spacer is an insertion into what was the 23S rRNA in prokaryotes.[26] The 45S rDNA is organized into 5 clusters (each has 30–40 repeats) on chromosomes 13, 14, 15, 21, and 22. These are transcribed by RNA polymerase I. The DNA for the 5S subunit occurs in tandem arrays (~200–300 true 5S genes and many dispersed pseudogenes), the largest one on the chromosome 1q41-42. 5S rRNA is transcribed by RNA polymerase III. The 18S rRNA in most eukaryotes is in the small ribosomal subunit, and the large subunit contains three rRNA species (the 5S, 5.8S and 28S in mammals, 25S in plants, rRNAs).
In flies, the large subunit contains four rRNA species instead of three with a split in the 5.8S rRNA that presents a shorter 5.8S subunit (123 nt) and a 30 nucleotide subunit named the 2S rRNA. Both fragments are separated by an internally transcribed spacer of 28 nucleotides. Since the 2S rRNA is small and highly abundant, its presence can interfere with construction of sRNA libraries and compromise the quantification of other sRNAs. The 2S subunit is retrieved in fruit fly and dark-winged fungus gnat species but absent from mosquitoes.[30]
The tertiary structure of the small subunit ribosomal RNA (SSU rRNA) has been resolved by X-ray crystallography.[31] The secondary structure of SSU rRNA contains 4 distinct domains—the 5', central, 3' major and 3' minor domains. A model of the secondary structure for the 5' domain (500-800 nucleotides) is shown.
Biosynthesis
In eukaryotes
As the building-blocks for the organelle, production of rRNA is ultimately the rate-limiting step in the synthesis of a ribosome. In the nucleolus, rRNA is synthesized by RNA polymerase I using the specialty genes (rDNA) that encode for it, which are found repeatedly throughout the genome.[32] The genes coding for 18S, 28S and 5.8S rRNA are located in the nucleolus organizer region and are transcribed into large precursor rRNA (pre-rRNA) molecules by RNA polymerase I. These pre-rRNA molecules are separated by external and internal spacer sequences and then methylated, which is key for later assembly and folding.[33][34][35] After separation and release as individual molecules, assembly proteins bind to each naked rRNA strand and fold it into its functional form using cooperative assembly and progressive addition of more folding proteins as needed. The exact details of how the folding proteins bind to the rRNA and how correct folding is achieved remains unknown.[36] The rRNA complexes are then further processed by reactions involving exo- and endo-nucleolytic cleavages guided by snoRNA (small nucleolar RNAs) in complex with proteins. As these complexes are compacted together to form a cohesive unit, interactions between rRNA and surrounding ribosomal proteins are constantly remodeled throughout assembly in order to provide stability and protect binding sites.[37] This process is referred to as the "maturation" phase of the rRNA lifecycle. The modifications that occur during maturation of rRNA have been found to contribute directly to control of gene expression by providing physical regulation of translational access of tRNA and mRNA.[38] Some studies have found that extensive methylation of various rRNA types is also necessary during this time to maintain ribosome stability.[39][40]
The genes for 5S rRNA are located inside the nucleolus and are transcribed into pre-5S rRNA by RNA polymerase III.[41] The pre-5S rRNA enters the nucleolus for processing and assembly with 28S and 5.8S rRNA to form the LSU. 18S rRNA forms the SSUs by combining with numerous ribosomal proteins. Once both subunits are assembled, they are individually exported into the cytoplasm to form the 80S unit and begin initiation of translation of mRNA.[42][43]
Ribosomal RNA is non-coding and is never translated into proteins of any kind: rRNA is only transcribed from rDNA and then matured for use as a structural building block for ribosomes. Transcribed rRNA is bound to ribosomal proteins to form the subunits of ribosomes and acts as the physical structure that pushes mRNA and tRNA through the ribosome to process and translate them.[1]
Eukaryotic regulation
Synthesis of rRNA is up-regulated and down-regulated to maintain homeostasis by a variety of processes and interactions:
- The kinase AKT indirectly promotes synthesis of rRNA as RNA polymerase I is AKT-dependent.[44]
- Certain angiogenic ribonucleases, such as angiogenin (ANG), can translocate and accumulate in the nucleolus. When the concentration of ANG becomes too high, some studies have found that ANG can bind to the promoter region of rDNA and unnecessarily increase rRNA transcription. This can be damaging to the nucleolus and can even lead to unchecked transcription and cancer.[45]
- During times of cellular glucose restriction, AMP-activated protein kinase (AMPK) discourages metabolic processes that consume energy but are non-essential. As a result, it is capable of phosphorylating RNA polymerase I (at the Ser-635 site) in order to down-regulate rRNA synthesis by disrupting transcription initiation.[46]
- Impairment or removal of more than one pseudouridine or 29-O-methylation regions from the ribosome decoding center significantly reduces rate of rRNA transcription by reducing the rate of incorporation of new amino acids.[47]
- Formation of heterochromatin is essential to silencing rRNA transcription, without which ribosomal RNA is synthesized unchecked and greatly decreases the lifespan of the organism.[48]
In prokaryotes
Similar to eukaryotes, the production of rRNA is the rate-limiting step in the prokaryotic synthesis of a ribosome. In E. coli, it has been found that rRNA is transcribed from the two promoters P1 and P2 found within seven different rrn operons. The P1 promoter is specifically responsible for regulating rRNA synthesis during moderate to high bacterial growth rates. Because the transcriptional activity of this promoter is directly proportional to the growth rate, it is primarily responsible for rRNA regulation. An increased rRNA concentration serves as a negative feedback mechanism to ribosome synthesis. High NTP concentration has been found to be required for efficient transcription of the rrn P1 promoters. They are thought to form stabilizing complexes with RNA polymerase and the promoters. In bacteria specifically, this association of high NTP concentration with increased rRNA synthesis provides a molecular explanation as to why ribosomal and thus protein synthesis is dependent on growth-rate. A low growth-rate yields lower rRNA / ribosomal synthesis rates while a higher growth rate yields a higher rRNA / ribosomal synthesis rate. This allows a cell to save energy or increase its metabolic activity dependent on its needs and available resources.[49][50][51]
In prokaryotic cells, each rRNA gene or operon is transcribed into a single RNA precursor that includes 16S, 23S, 5S rRNA and tRNA sequences along with transcribed spacers. The RNA processing then begins before the transcription is complete. During processing reactions, the rRNAs and tRNAs are released as separate molecules.[52]
Prokaryotic regulation
Because of the vital role rRNA plays in the cell physiology of prokaryotes, there is much overlap in rRNA regulation mechanisms. At the transcriptional level, there are both positive and negative effectors of rRNA transcription that facilitate a cell's maintenance of homeostasis:
- An UP element upstream of the rrn P1 promoter can bind a subunit of RNA polymerase, thus promoting transcription of rRNA.
- Transcription factors such as FIS bind upstream of the promoter and interact with RNA polymerase which facilitates transcription.
- Anti-termination factors bind downstream of the rrn P2 promoter, preventing premature transcription termination.
- Due to the stringent response, when the availability of amino acids is low, ppGpp (a negative effector) can inhibit transcription from both the P1 and P2 promoters.[49]
Degradation
Ribosomal RNA is quite stable in comparison to other common types of RNA and persists for longer periods of time in a healthy cellular environment. Once assembled into functional units, ribosomal RNA within ribosomes are stable in the stationary phase of the cell life cycle for many hours.[53] Degradation can be triggered via "stalling" of a ribosome, a state that occurs when the ribosome recognizes faulty mRNA or encounters other processing difficulties that causes translation by the ribosome to cease. Once a ribosome stalls, a specialized pathway on the ribosome is initiated to target the entire complex for disassembly.[54]
In eukaryotes
As with any protein or RNA, rRNA production is prone to errors resulting in the production of non-functional rRNA. To correct this, the cell allows for degradation of rRNA through the non-functional rRNA decay (NRD) pathway.[55] Much of the research in this topic was conducted on eukaryotic cells, specifically Saccharomyces cerevisiae yeast. Currently, only a basic understanding of how cells are able to target functionally defective ribosomes for ubiquination and degradation in eukaryotes is available.[56]
- The NRD pathway for the 40S subunit may be independent or separate from the NRD pathway for the 60S subunit. It has been observed that certain genes were able to affect degradation of certain pre-RNAs, but not others.[57]
- Numerous proteins are involved in the NRD pathway, such as Mms1p and Rtt101p, which are believed to complex together to target ribosomes for degradation. Mms1p and Rtt101p are found to bind together and Rtt101p is believed to recruit a ubiquitin E3 ligase complex, allowing for the non-functional ribosomes to be ubiquinated before being degraded.[58]
- Prokaryotes lack a homolog for Mms1, so it is unclear how prokaryotes are able to degrade non-functional rRNAs.
- The growth rate of eukaryotic cells did not seem to be significantly affected by the accumulation of non-functional rRNAs.
In prokaryotes
Although there is far less research available on ribosomal RNA degradation in prokaryotes in comparison to eukaryotes, there has still been interest on whether bacteria follow a similar degradation scheme in comparison to the NRD in eukaryotes. Much of the research done for prokaryotes has been conducted on Escherichia coli. Many differences were found between eukaryotic and prokaryotic rRNA degradation, leading researchers to believe that the two degrade using different pathways.[59]
- Certain mutations in rRNA that were able to trigger rRNA degradation in eukaryotes were unable to do so in prokaryotes.
- Point mutations in a 23S rRNA would cause both 23S and 16S rRNAs to be degraded, in comparison to eukaryotes, in which mutations in one subunit would only cause that subunit to be degraded.
- Researchers found that removal of a whole helix structure (H69) from the 23S rRNA did not trigger its degradation. This led them to believe that H69 was critical for endonucleases to recognize and degrade the mutated rRNA.
Sequence conservation and stability
Due to the prevalent and unwavering nature of rRNA across all organisms, the study of its resistance to gene transfer, mutation, and alteration without destruction of the organism has become a popular field of interest. Ribosomal RNA genes have been found to be tolerant to modification and incursion. When rRNA sequencing is altered, cells have been found to become compromised and quickly cease normal function.[60] These key traits of rRNA have become especially important for gene database projects (comprehensive online resources such as SILVA[61] or SINA[62]) where alignment of ribosomal RNA sequences from across the different biologic domains greatly eases "taxonomic assignment, phylogenetic analysis and the investigation of microbial diversity."[61]
Examples of resilience:
- Addition of large, nonsensical RNA fragments into many parts of the 16S rRNA unit does not observably alter the function of the ribosomal unit as a whole.[63]
- Non-coding RNARD7 has the capability to alter processing of rRNA to make the molecules resistant to degradation by carboxylic acid. This is a crucial mechanism in maintaining rRNA concentrations during active growth when acid build-up (due to the substrate phosphorylation required to produce ATP) can become toxic to intracellular functions.[64]
- Insertion of hammerhead ribozymes that are capable of cis-cleavages along 16S rRNA greatly inhibit function and diminish stability.[63]
- While most cellular functions degrade heavily after only short period of exposure to hypoxic environments, rRNA remains un-degraded and resolved after six days of prolonged hypoxia. Only after such an extended period of time do rRNA intermediates (indicative of degradation finally occurring) begin to present themselves.[65]
Significance
Ribosomal RNA characteristics are important in evolution, thus taxonomy and medicine.
- rRNA is one of only a few gene products present in all cells.[43] For this reason, genes that encode the rRNA (rDNA) are sequenced to identify an organism's taxonomic group, calculate related groups, and estimate rates of species divergence.[66] As a result, many thousands of rRNA sequences are known and stored in specialized databases such as RDP-II[67] and SILVA.[68]
- Alterations to rRNA are what allow certain disease-causing bacteria, such as Mycobacterium tuberculosis (the bacterium that causes tuberculosis) to develop extreme drug resistance.[69] Due to similar issues, this has become a prevalent problem in veterinary medicine where the main method for handling bacterial infection in pets is administration of drugs that attack the peptidyl-transferase centre (PTC) of the bacterial ribosome. Mutations in 23S rRNA have created perfect resistance to these drugs as they operate together in an unknown fashion to bypass the PTC entirely.[70]
- rRNA is the target of numerous clinically relevant antibiotics: chloramphenicol, erythromycin, kasugamycin, micrococcin, paromomycin, ricin, alpha-sarcin, spectinomycin, streptomycin, and thiostrepton.
- rRNA have been shown to be the origin of species-specific microRNAs, like miR-663 in humans and miR-712 in mice. These particular miRNAs originate from the internal transcribed spacers of the rRNA.[71]
Human genes
- 45S: RNR1, RNR2, RNR3, RNR4, RNR5; (unclustered) RNA18SN1, RNA18SN2, RNA18SN3, RNA18SN4, RNA18SN5, RNA28SN1, RNA28SN2, RNA28SN3, RNA28SN4, RNA28SN5, RNA45SN1, RNA45SN2, RNA45SN3, RNA45SN4, RNA45SN5, RNA5-8SN1, RNA5-8SN2, RNA5-8SN3, RNA5-8SN4, RNA5-8SN5
- 5S: RNA5S1, RNA5S2, RNA5S3, RNA5S4, RNA5S5, RNA5S6, RNA5S7, RNA5S8, RNA5S9, RNA5S10, RNA5S11, RNA5S12, RNA5S13, RNA5S14, RNA5S15, RNA5S16, RNA5S17
- Mt: MT-RNR1, MT-TV (co-opted), MT-RNR2
See also
- Ribotyping
- Diazaborine B, a maturation inhibitor of rRNAs for the large ribosomal subunit
References
- 1 2 Berk, Arnold; Baltimore, David; Lodish, Harvey; Darnell, James; Matsudaira, Paul; Zipursky, S. Lawrence (1996-01-31). Molekulare Zellbiologie. Berlin, Boston: DE GRUYTER. doi:10.1515/9783110810578. ISBN 9783110810578.
- ↑ Lodish, Harvey; Berk, Arnold; Zipursky, S. Lawrence; Matsudaira, Paul; Baltimore, David; Darnell, James (2000). "The Three Roles of RNA in Protein Synthesis". Molecular Cell Biology. 4th Edition.
- ↑ Urlaub H, Kruft V, Bischof O, Müller EC, Wittmann-Liebold B (September 1995). "Protein-rRNA binding features and their structural and functional implications in ribosomes as determined by cross-linking studies". The EMBO Journal. 14 (18): 4578–88. doi:10.1002/j.1460-2075.1995.tb00137.x. PMC 394550. PMID 7556101.
- ↑ Ferreira-Cerca S, Pöll G, Gleizes PE, Tschochner H, Milkereit P (October 2005). "Roles of eukaryotic ribosomal proteins in maturation and transport of pre-18S rRNA and ribosome function". Molecular Cell. 20 (2): 263–75. doi:10.1016/j.molcel.2005.09.005. PMID 16246728.
- ↑ Szymański M, Barciszewska MZ, Erdmann VA, Barciszewski J (May 2003). "5 S rRNA: structure and interactions". The Biochemical Journal. 371 (Pt 3): 641–51. doi:10.1042/bj20020872. PMC 1223345. PMID 12564956.
- ↑ Henras AK, Plisson-Chastang C, O'Donohue MF, Chakraborty A, Gleizes PE (2015-03-01). "An overview of pre-ribosomal RNA processing in eukaryotes". Wiley Interdisciplinary Reviews: RNA. 6 (2): 225–42. doi:10.1002/wrna.1269. PMC 4361047. PMID 25346433.
- ↑ Penev PI, Fakhretaha-Aval S, Patel VJ, Cannone JJ, Gutell RR, Petrov AS, Williams LD, Glass JB (August 2020). "Supersized ribosomal RNA expansion segments in Asgard archaea". Genome Biology and Evolution. 12 (10): 1694–1710. doi:10.1093/gbe/evaa170. PMC 7594248. PMID 32785681.
- ↑ Luehrsen, KR.; Nicholson, DE; Eubanks, DC; Fox, GE (May 1981). "An archaebacterial 5S rRNA contains a long insertion sequence". Nature. 293 (5835): 755–756. Bibcode:1981Natur.293..755L. doi:10.1038/293755a0. PMID 6169998. S2CID 4341755.
- ↑ Tirumalai, MR; Kaelber, JT; Park, DR; Tran, Q; Fox, GE (31 August 2020). "Cryo-electron microscopy visualization of a large insertion in the 5S ribosomal RNA of the extremely halophilic archaeon Halococcus morrhuae". FEBS Open Bio. 10 (10): 1938–1946. doi:10.1002/2211-5463.12962. PMC 7530397. PMID 32865340.
- ↑ Woese CR, Fox GE (November 1977). "Phylogenetic structure of the prokaryotic domain: the primary kingdoms". Proc. Natl. Acad. Sci. USA. 74 (11): 5088–5090. Bibcode:1977PNAS...74.5088W. doi:10.1073/pnas.74.11.5088. PMC 432104. PMID 270744.
- ↑ Lagesen K, Hallin P, Rødland EA, Staerfeldt HH, Rognes T, Ussery DW (2007-05-01). "RNAmmer: consistent and rapid annotation of ribosomal RNA genes". Nucleic Acids Research. 35 (9): 3100–8. doi:10.1093/nar/gkm160. PMC 1888812. PMID 17452365.
- ↑ Chun J, Lee JH, Jung Y, Kim M, Kim S, Kim BK, Lim YW (October 2007). "EzTaxon: a web-based tool for the identification of prokaryotes based on 16S ribosomal RNA gene sequences". International Journal of Systematic and Evolutionary Microbiology. 57 (Pt 10): 2259–61. doi:10.1099/ijs.0.64915-0. PMID 17911292.
- 1 2 3 Tirumalai MR, Rivas M, Tran Q, Fox GE (November 2021). "The Peptidyl Transferase Center: a Window to the Past". Microbiol Mol Biol Rev. 85 (4): e0010421. doi:10.1128/MMBR.00104-21. PMC 8579967. PMID 34756086.
- ↑ Ghosh, Arnab; Komar, Anton A (2 January 2015). "Eukaryote-specific extensions in ribosomal proteins of the small subunit: Structure and function". Translation. 3 (1): e999576. doi:10.1080/21690731.2014.999576. PMC 4682806. PMID 26779416.
- ↑ Lodmell JS, Dahlberg AE (August 1997). "A conformational switch in Escherichia coli 16S ribosomal RNA during decoding of messenger RNA". Science. 277 (5330): 1262–7. doi:10.1126/science.277.5330.1262. PMID 9271564.
- ↑ Gabashvili IS, Agrawal RK, Grassucci R, Squires CL, Dahlberg AE, Frank J (November 1999). "Major rearrangements in the 70S ribosomal 3D structure caused by a conformational switch in 16S ribosomal RNA". The EMBO Journal. 18 (22): 6501–7. doi:10.1093/emboj/18.22.6501. PMC 1171713. PMID 10562562.
- ↑ Woolford JL, Baserga SJ (November 2013). "Ribosome biogenesis in the yeast Saccharomyces cerevisiae". Genetics. 195 (3): 643–81. doi:10.1534/genetics.113.153197. PMC 3813855. PMID 24190922.
- 1 2 Baßler J, Hurt E (June 2019). "Eukaryotic Ribosome Assembly". Annual Review of Biochemistry. 88 (1): 281–306. doi:10.1146/annurev-biochem-013118-110817. PMID 30566372. S2CID 58650367.
- ↑ Moore PB, Steitz TA (July 2002). "The involvement of RNA in ribosome function". Nature. 418 (6894): 229–35. Bibcode:2002Natur.418..229M. doi:10.1038/418229a. PMID 12110899. S2CID 4324362.
- ↑ "Homo sapiens RNA, 5S ribosomal". National Center for Biotechnology Information (NCBI). 2020-09-03. Retrieved 2024-01-06.
- ↑ "Homo sapiens 5.8S ribosomal RNA". National Center for Biotechnology Information (NCBI). 2017-02-10.
- ↑ "Homo sapiens 28S ribosomal RNA". National Center for Biotechnology Information (NCBI). 2017-02-04.
- ↑ "Homo sapiens 18S ribosomal RNA". National Center for Biotechnology Information (NCBI). 2017-02-04.
- ↑ Kaushal, PS; Sharma, MR; Agrawal, RK (July 2015). "The 55S mammalian mitochondrial ribosome and its tRNA-exit region". Biochimie. 114: 119–26. doi:10.1016/j.biochi.2015.03.013. PMC 4772884. PMID 25797916.
- 1 2 3 Stoddard SF, Smith BJ, Hein R, Roller BR, Schmidt TM (January 2015). "rrnDB: improved tools for interpreting rRNA gene abundance in bacteria and archaea and a new foundation for future development". Nucleic Acids Research. 43 (Database issue): D593-8. doi:10.1093/nar/gku1201. PMC 4383981. PMID 25414355.
- 1 2 Lafontaine DL, Tollervey D (July 2001). "The function and synthesis of ribosomes". Nature Reviews Molecular Cell Biology. 2 (7): 514–20. doi:10.1038/35080045. hdl:1842/729. PMID 11433365. S2CID 2637106.
- ↑ Stults DM, Killen MW, Williamson EP, Hourigan JS, Vargas HD, Arnold SM, et al. (December 2009). "Human rRNA gene clusters are recombinational hotspots in cancer". Cancer Research. 69 (23): 9096–104. doi:10.1158/0008-5472.can-09-2680. PMID 19920195. S2CID 6162867.
- ↑ Kim JH, Dilthey AT, Nagaraja R, Lee HS, Koren S, Dudekula D, et al. (July 2018). "Variation in human chromosome 21 ribosomal RNA genes characterized by TAR cloning and long-read sequencing". Nucleic Acids Research. 46 (13): 6712–6725. doi:10.1093/nar/gky442. PMC 6061828. PMID 29788454.
- ↑ Parks MM, Kurylo CM, Dass RA, Bojmar L, Lyden D, Vincent CT, Blanchard SC (February 2018). "Variant ribosomal RNA alleles are conserved and exhibit tissue-specific expression". Science Advances. 4 (2): eaao0665. Bibcode:2018SciA....4..665P. doi:10.1126/sciadv.aao0665. PMC 5829973. PMID 29503865.
- ↑ Shimada, T. (August 1992). "Distribution of split 5.8S ribosomal RNA in Diptera". Insect Molecular Biology. 1 (1): 45–48. doi:10.1111/j.1365-2583.1993.tb00076.x. ISSN 0962-1075. PMID 1343775. S2CID 46570307.
- ↑ Yusupov MM, Yusupova GZ, Baucom A, Lieberman K, Earnest TN, Cate JH, Noller HF (May 2001). "Crystal structure of the ribosome at 5.5 A resolution". Science. 292 (5518): 883–96. Bibcode:2001Sci...292..883Y. doi:10.1126/science.1060089. PMID 11283358. S2CID 39505192.
- ↑ "Ribosomal RNA | genetics". Encyclopedia Britannica. Retrieved 2019-10-02.
- ↑ Zemora G, Waldsich C (November 2010). "RNA folding in living cells". RNA Biology. 7 (6): 634–41. doi:10.4161/rna.7.6.13554. PMC 3073324. PMID 21045541.
- ↑ Fernández-Tornero C, Moreno-Morcillo M, Rashid UJ, Taylor NM, Ruiz FM, Gruene T, et al. (October 2013). "Crystal structure of the 14-subunit RNA polymerase I". Nature. 502 (7473): 644–9. Bibcode:2013Natur.502..644F. doi:10.1038/nature12636. PMID 24153184. S2CID 205235881.
- ↑ Engel C, Sainsbury S, Cheung AC, Kostrewa D, Cramer P (October 2013). "RNA polymerase I structure and transcription regulation". Nature. 502 (7473): 650–5. Bibcode:2013Natur.502..650E. doi:10.1038/nature12712. hdl:11858/00-001M-0000-0015-3B48-5. PMID 24153182. S2CID 205236187.
- ↑ Dutca LM, Gallagher JE, Baserga SJ (July 2011). "The initial U3 snoRNA:pre-rRNA base pairing interaction required for pre-18S rRNA folding revealed by in vivo chemical probing". Nucleic Acids Research. 39 (12): 5164–80. doi:10.1093/nar/gkr044. PMC 3130255. PMID 21349877.
- ↑ Woodson SA (December 2011). "RNA folding pathways and the self-assembly of ribosomes". Accounts of Chemical Research. 44 (12): 1312–9. doi:10.1021/ar2000474. PMC 4361232. PMID 21714483.
- ↑ Sloan KE, Warda AS, Sharma S, Entian KD, Lafontaine DL, Bohnsack MT (September 2017). "Tuning the ribosome: The influence of rRNA modification on eukaryotic ribosome biogenesis and function". RNA Biology. 14 (9): 1138–1152. doi:10.1080/15476286.2016.1259781. PMC 5699541. PMID 27911188.
- ↑ Gigova A, Duggimpudi S, Pollex T, Schaefer M, Koš M (October 2014). "A cluster of methylations in the domain IV of 25S rRNA is required for ribosome stability". RNA. 20 (10): 1632–44. doi:10.1261/rna.043398.113. PMC 4174444. PMID 25125595.
- ↑ Metodiev MD, Lesko N, Park CB, Cámara Y, Shi Y, Wibom R, et al. (April 2009). "Methylation of 12S rRNA is necessary for in vivo stability of the small subunit of the mammalian mitochondrial ribosome". Cell Metabolism. 9 (4): 386–97. doi:10.1016/j.cmet.2009.03.001. PMID 19356719.
- ↑ Thompson M, Haeusler RA, Good PD, Engelke DR (November 2003). "Nucleolar clustering of dispersed tRNA genes". Science. 302 (5649): 1399–401. Bibcode:2003Sci...302.1399T. doi:10.1126/science.1089814. PMC 3783965. PMID 14631041.
- ↑ "rRNA synthesis and processing".
- 1 2 Smit S, Widmann J, Knight R (2007). "Evolutionary rates vary among rRNA structural elements". Nucleic Acids Research. 35 (10): 3339–54. doi:10.1093/nar/gkm101. PMC 1904297. PMID 17468501.
- ↑ Chan JC, Hannan KM, Riddell K, Ng PY, Peck A, Lee RS, et al. (August 2011). "AKT promotes rRNA synthesis and cooperates with c-MYC to stimulate ribosome biogenesis in cancer". Science Signaling. 4 (188): ra56. doi:10.1126/scisignal.2001754. PMID 21878679. S2CID 20979505.
- ↑ Li S, Ibaragi S, Hu GF (May 2011). "Angiogenin as a molecular target for the treatment of prostate cancer". Current Cancer Therapy Reviews. 7 (2): 83–90. doi:10.2174/1573394711107020083. PMC 3131147. PMID 21743803.
- ↑ Hoppe S, Bierhoff H, Cado I, Weber A, Tiebe M, Grummt I, Voit R (October 2009). "AMP-activated protein kinase adapts rRNA synthesis to cellular energy supply". Proceedings of the National Academy of Sciences of the United States of America. 106 (42): 17781–6. Bibcode:2009PNAS..10617781H. doi:10.1073/pnas.0909873106. PMC 2764937. PMID 19815529.
- ↑ Liang XH, Liu Q, Fournier MJ (September 2009). "Loss of rRNA modifications in the decoding center of the ribosome impairs translation and strongly delays pre-rRNA processing". RNA. 15 (9): 1716–28. doi:10.1261/rna.1724409. PMC 2743053. PMID 19628622.
- ↑ Larson K, Yan SJ, Tsurumi A, Liu J, Zhou J, Gaur K, et al. (January 2012). "Heterochromatin formation promotes longevity and represses ribosomal RNA synthesis". PLOS Genetics. 8 (1): e1002473. doi:10.1371/journal.pgen.1002473. PMC 3266895. PMID 22291607.
- 1 2 Gaal T, Bartlett MS, Ross W, Turnbough CL, Gourse RL (December 1997). "Transcription regulation by initiating NTP concentration: rRNA synthesis in bacteria". Science. 278 (5346): 2092–7. Bibcode:1997Sci...278.2092G. doi:10.1126/science.278.5346.2092. PMID 9405339.
- ↑ Maeda M, Shimada T, Ishihama A (2015-12-30). "Strength and Regulation of Seven rRNA Promoters in Escherichia coli". PLOS ONE. 10 (12): e0144697. Bibcode:2015PLoSO..1044697M. doi:10.1371/journal.pone.0144697. PMC 4696680. PMID 26717514.
- ↑ Gaal T, Bratton BP, Sanchez-Vazquez P, Sliwicki A, Sliwicki K, Vegel A, et al. (October 2016). "Colocalization of distant chromosomal loci in space in E. coli: a bacterial nucleolus". Genes & Development. 30 (20): 2272–2285. doi:10.1101/gad.290312.116. PMC 5110994. PMID 27898392.
- ↑ Wolfe, Stephen (1993). Molecular and Cellular Biology. Wadsworth Publishing Company. ISBN 978-0534124083.
- ↑ Piir K, Paier A, Liiv A, Tenson T, Maiväli U (May 2011). "Ribosome degradation in growing bacteria". EMBO Reports. 12 (5): 458–62. doi:10.1038/embor.2011.47. PMC 3090016. PMID 21460796.
- ↑ Brandman O, Hegde RS (January 2016). "Ribosome-associated protein quality control". Nature Structural & Molecular Biology. 23 (1): 7–15. doi:10.1038/nsmb.3147. PMC 4853245. PMID 26733220.
- ↑ Fujii K, Kitabatake M, Sakata T, Miyata A, Ohno M (April 2009). "A role for ubiquitin in the clearance of nonfunctional rRNAs". Genes & Development. 23 (8): 963–74. doi:10.1101/gad.1775609. PMC 2675866. PMID 19390089.
- ↑ Donovan, Bridget M.; Jarrell, Kelli L.; LaRiviere, Frederick J. (2011-04-01). "Investigating nonfunctional rRNA decay as a stress response in Saccharomyces cerevisiae". The FASEB Journal. 25 (1_supplement): 521.3. doi:10.1096/fasebj.25.1_supplement.521.3 (inactive 1 August 2023). ISSN 0892-6638.
{{cite journal}}: CS1 maint: DOI inactive as of August 2023 (link) - ↑ LaRiviere FJ, Cole SE, Ferullo DJ, Moore MJ (November 2006). "A late-acting quality control process for mature eukaryotic rRNAs". Molecular Cell. 24 (4): 619–26. doi:10.1016/j.molcel.2006.10.008. PMID 17188037.
- ↑ Michel JJ, McCarville JF, Xiong Y (June 2003). "A role for Saccharomyces cerevisiae Cul8 ubiquitin ligase in proper anaphase progression". The Journal of Biological Chemistry. 278 (25): 22828–37. doi:10.1074/jbc.M210358200. PMID 12676951. S2CID 33099674.
- ↑ Paier A, Leppik M, Soosaar A, Tenson T, Maiväli Ü (January 2015). "The effects of disruptions in ribosomal active sites and in intersubunit contacts on ribosomal degradation in Escherichia coli". Scientific Reports. 5: 7712. Bibcode:2015NatSR...5E7712P. doi:10.1038/srep07712. PMC 4289901. PMID 25578614.
- ↑ Ide S, Miyazaki T, Maki H, Kobayashi T (February 2010). "Abundance of ribosomal RNA gene copies maintains genome integrity". Science. 327 (5966): 693–6. Bibcode:2010Sci...327..693I. doi:10.1126/science.1179044. PMID 20133573. S2CID 206522454.
- 1 2 Quast C, Pruesse E, Yilmaz P, Gerken J, Schweer T, Yarza P, et al. (January 2013). "The SILVA ribosomal RNA gene database project: improved data processing and web-based tools". Nucleic Acids Research. 41 (Database issue): D590-6. doi:10.1093/nar/gks1219. PMC 3531112. PMID 23193283.
- ↑ Pruesse E, Peplies J, Glöckner FO (July 2012). "SINA: accurate high-throughput multiple sequence alignment of ribosomal RNA genes". Bioinformatics. 28 (14): 1823–9. doi:10.1093/bioinformatics/bts252. PMC 3389763. PMID 22556368.
- 1 2 Wieland M, Berschneider B, Erlacher MD, Hartig JS (March 2010). "Aptazyme-mediated regulation of 16S ribosomal RNA". Chemistry & Biology. 17 (3): 236–42. doi:10.1016/j.chembiol.2010.02.012. PMID 20338515.
- ↑ Borden JR, Jones SW, Indurthi D, Chen Y, Papoutsakis ET (May 2010). "A genomic-library based discovery of a novel, possibly synthetic, acid-tolerance mechanism in Clostridium acetobutylicum involving non-coding RNAs and ribosomal RNA processing". Metabolic Engineering. 12 (3): 268–81. doi:10.1016/j.ymben.2009.12.004. PMC 2857598. PMID 20060060.
- ↑ Trauner A, Lougheed KE, Bennett MH, Hingley-Wilson SM, Williams HD (July 2012). "The dormancy regulator DosR controls ribosome stability in hypoxic mycobacteria". The Journal of Biological Chemistry. 287 (28): 24053–63. doi:10.1074/jbc.m112.364851. PMC 3390679. PMID 22544737.
- ↑ Meyer A, Todt C, Mikkelsen NT, Lieb B (March 2010). "Fast evolving 18S rRNA sequences from Solenogastres (Mollusca) resist standard PCR amplification and give new insights into mollusk substitution rate heterogeneity". BMC Evolutionary Biology. 10 (1): 70. doi:10.1186/1471-2148-10-70. PMC 2841657. PMID 20214780.
- ↑ Cole JR, Chai B, Marsh TL, Farris RJ, Wang Q, Kulam SA, et al. (January 2003). "The Ribosomal Database Project (RDP-II): previewing a new autoaligner that allows regular updates and the new prokaryotic taxonomy". Nucleic Acids Research. 31 (1): 442–3. doi:10.1093/nar/gkg039. PMC 165486. PMID 12520046.
- ↑ Pruesse E, Quast C, Knittel K, Fuchs BM, Ludwig W, Peplies J, Glöckner FO (2007). "SILVA: a comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB". Nucleic Acids Research. 35 (21): 7188–96. doi:10.1093/nar/gkm864. PMC 2175337. PMID 17947321.
- ↑ Wade, M.; Zhang, Y. (2005), "Mechanisms of Drug Resistance in Mycobacterium tuberculosis", Tuberculosis and the Tubercle Bacillus, American Society of Microbiology, pp. 115–140, doi:10.1128/9781555817657.ch8, ISBN 9781555817657, S2CID 36002898
- ↑ Long KS, Poehlsgaard J, Hansen LH, Hobbie SN, Böttger EC, Vester B (March 2009). "Single 23S rRNA mutations at the ribosomal peptidyl transferase centre confer resistance to valnemulin and other antibiotics in Mycobacterium smegmatis by perturbation of the drug binding pocket". Molecular Microbiology. 71 (5): 1218–27. doi:10.1111/j.1365-2958.2009.06596.x. PMID 19154331. S2CID 23728518.
- ↑ Ju Son D (2013). "The atypical mechanosensitive microRNA-712 derived from pre-ribosomal RNA induces endothelial inflammation and atherosclerosis". Nature Communications. 4: 3000. Bibcode:2013NatCo...4.3000S. doi:10.1038/ncomms4000. PMC 3923891. PMID 24346612.
External links
- 16S rRNA, BioMineWiki Archived 2019-04-27 at the Wayback Machine
- Ribosomal Database Project II Archived 2020-08-19 at the Wayback Machine
- Ribosomal+RNA at the U.S. National Library of Medicine Medical Subject Headings (MeSH)
- SILVA rRNA Database Project (also includes Eukaryotes (18S) and LSU (23S/28S))
- Video: rRNA: sequence, function & synthesis
- Halococcus morrhuae (archaebacterium) 5S rRNA
Types of nucleic acids | |||||||
|---|---|---|---|---|---|---|---|
| Constituents | |||||||
| Ribonucleic acids (coding, non-coding) |
| ||||||
| Deoxyribonucleic acids | |||||||
| Analogues | |||||||
| Cloning vectors | |||||||
| |||||||