| S100A7A | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Identifiers | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Aliases | S100A7A, NICE-2, S100A15, S100A7L1, S100A7f, S100 calcium binding protein A7A, NICE2 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| External IDs | OMIM: 617427 GeneCards: S100A7A | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Wikidata | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||

Protein S100-A7A (S100A7A), also known as koebnerisin, is a protein that in humans is encoded by the S100A7A (alias: S100A15) gene.[3]
S100 proteins are a diverse calcium-binding family that regulate fundamental cellular and extracellular processes including cell proliferation and differentiation, cell migration, and the antimicrobial host defense as antimicrobial peptides.
Koebnerisin (S100A7A) was first identified upregulated in inflammation-prone psoriatic skin, suggesting involvement in the lesional phenotype of the disease,[4] Koebner phenomenon. Today, the protein is of further interest because of its role in antimicrobial defence, innate immunity, epidermal cell maturation and epithelial tumorigenesis.[5][6]
Function
Epithelial homeostasis and antimicrobial host defense
Skin: In normal epidermis, koebnerisin (S100A7A) is expressed by epidermal basal and differentiated keratinocytes, melanocytes, and Langerhans cells. Within the pilosebaceous unit, S100A7A is found in the inner and external root sheath and the basal layer of the sebaceous gland. In the dermis, koebnerisin (S100A7A) is produced by dendritic cells, smooth muscle cells, endothelial cells, as well as fibroblasts to control tissue regeneration.[7][8][9][10]
Breast: Koebnerisin (S100A7A) is expressed by alveolar and small duct luminal cells and by epithelial-derived myoepithelial cells around acini and by surrounding blood vessels.[11]
Koebnerisin (S100A7A) functions as an antimicrobial peptide (AMP) reducing survival of E. coli and was strongly regulated by several bacterial components, such as Pseudomonas aeruginosa and Staphylococcus aureus. Thus, koebnerisin participates in the antimicrobial host defence of the skin and in the digestive tract of breast-feeding newborns.[12]
Epithelial carcinogenesis
Breast cancer: Koebnerisin (S100A7A) is overexpressed in ER/PR negative tumors suggesting a regulation with tumor progression.[11] The secreted koebnerisin (S100A7A) acts as a chemoattractant,[8] enhances inflammation and thus could drive breast carcinogenesis.
Inflammation
Koebnerisin (S100A7A) is overexpressed in inflammatory skin diseases, such as psoriasis and eczema.[13] It is regulated through Th1 and Th17 but not Th2 proinflammatory cytokines.[14][15] When released into the extracellar space, koebnerisin (S100A7A) induces inflammation. It acts as a chemoattractant for myeloid leukocytes through a pertussis toxin sensitive Gi protein coupled receptor. Koebnerisin (S100A7A) amplifies inflammation with related psoriasin (S100A7) that is co-regulated and proinflammatory through RAGE.
Genomic organization and mRNA splice variants
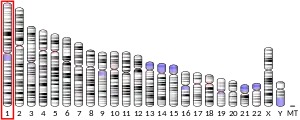
Koebnerisin (S100A7A) maps to the S100 gene cluster within the epidermal differentiation complex (EDC, chromosome 1q21) and reveals an unusual genomic organization compared to other S100 members. The two alternative mRNA-isoforms of koebnerisin share the same coding region, but show differences in composition and length of adjacent untranslated regions (S100A7A-short (S): 0.5 kb vs. hS100A7A-long (L): 4.4 kb). Both splice variants are differently regulated in inflammatory skin diseases suggesting usage of alternate promoters.[4][14]
Protein
The amino acid sequence reveals a conserved C-terminal and a variant N-terminal EF-hand typical for S100 proteins (101 amino acids, 11.305 Da, calculated pI of 7.57 kDa). Compared to most S100 proteins, koebnerisin (S100A7A) is basic.
Evolution
Primate
Koebnerisin (S100A7A) has lately evolved by gene duplications within the Epidermal Differentiation Complex (EDC, chromosome 1q21) during primate evolution forming a novel S100 subfamily together with Psoriasin (S100A7).[16][17] Therefore, koebnerisin is almost identical to psoriasin in sequence (>90%). Despite their high homology, koebnerisin (S100A7A) and psoriasin (S100A7) are distinct in tissue distribution, regulation, structure[18][19] and function and, thus exemplary for the diversity within the S100 family. Their different properties are compelling reasons to discriminate S100A7A (koebnerisin) and S100A7 (psoriasin) in epithelial homeostasis, inflammation and cancer.
Rodent
Koebnerisin (S100A7A) and psoriasin (S100A7) share a common protein in mice encoded by the S100a7a15 (alias: mS100A7, mS100A15, mS100a7a) gene.[20] It can be used to study the significance of the corresponding human proteins for epidermal maturation, inflammation and epithelial carcinogenesis.[17][21][22][23]
References
- 1 2 3 GRCh38: Ensembl release 89: ENSG00000184330 - Ensembl, May 2017
- ↑ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ↑ "Entrez Gene: S100 calcium binding protein A7A".
- 1 2 Wolf R, Mirmohammadsadegh A, Walz M, Lysa B, Tartler U, Remus R, Hengge U, Michel G, Ruzicka T (2003). "Molecular cloning and characterization of alternatively spliced mRNA isoforms from psoriatic skin encoding a novel member of the S100 family". FASEB J. 17 (13): 1969–71. doi:10.1096/fj.03-0148fje. PMID 12923069. S2CID 14817867.
- ↑ Wolf R, Ruzicka T, Yuspa SH (2011). "Novel S100A7 (psoriasin)/S100A15 (koebnerisin) subfamily: highly homologous but distinct in regulation and function". Amino Acids. 41 (4): 789–96. doi:10.1007/s00726-010-0666-4. PMC 6410564. PMID 20596736.
- ↑ Zwicker S, Bureik D, Ruzicka T, Wolf R (2012). "[Friend or Foe?--Psoriasin and Koebnerisin: multifunctional defence molecules in skin differentiation, tumorigenesis and inflammation]". Dtsch. Med. Wochenschr. (in German). 137 (10): 491–4. doi:10.1055/s-0031-1299015. PMID 22374659.
- ↑ Gauglitz GG, Bureik D, Zwicker S, Ruzicka T, Wolf R (2014). "The Antimicrobial Peptides Psoriasin (S100A7) and Koebnerisin (S100A15) Suppress Extracellular Matrix Production and Proliferation of Human Fibroblasts". Skin Pharmacol Physiol. 28 (3): 115–123. doi:10.1159/000363579. PMID 25502330. S2CID 23694258.
- 1 2 Wolf R, Howard OM, Dong HF, Voscopoulos C, Boeshans K, Winston J, Divi R, Gunsior M, Goldsmith P, Ahvazi B, Chavakis T, Oppenheim JJ, Yuspa SH (2008). "Chemotactic activity of S100A7 (Psoriasin) is mediated by the receptor for advanced glycation end products and potentiates inflammation with highly homologous but functionally distinct S100A15". J. Immunol. 181 (2): 1499–506. doi:10.4049/jimmunol.181.2.1499. PMC 2435511. PMID 18606705.
- ↑ Zhang X, Sharma AM, Uetrecht J (Sep 2013). "Identification of danger signals in nevirapine-induced skin rash". Chemical Research in Toxicology. 26 (9): 1378–83. doi:10.1021/tx400232s. PMID 23947594.
- ↑ de Castro A, Minty F, Hattinger E, Wolf R, Parkinson EK (2014). "The secreted protein S100A7 (psoriasin) is induced by telomere dysfunction in human keratinocytes independently of a DNA damage response and cell cycle regulators". Longevity & Healthspan. 3: 8. doi:10.1186/2046-2395-3-8. PMC 4304136. PMID 25621169.
- 1 2 Wolf R, Voscopoulos C, Winston J, Dharamsi A, Goldsmith P, Gunsior M, Vonderhaar BK, Olson M, Watson PH, Yuspa SH (2009). "Highly homologous hS100A15 and hS100A7 proteins are distinctly expressed in normal breast tissue and breast cancer". Cancer Lett. 277 (1): 101–7. doi:10.1016/j.canlet.2008.11.032. PMC 2680177. PMID 19136201.
- ↑ Büchau AS, Hassan M, Kukova G, Lewerenz V, Kellermann S, Würthner JU, Wolf R, Walz M, Gallo RL, Ruzicka T (2007). "S100A15, an antimicrobial protein of the skin: regulation by E. coli through Toll-like receptor 4". J. Invest. Dermatol. 127 (11): 2596–604. doi:10.1038/sj.jid.5700946. PMID 17625598.
- ↑ Wolf R, Mascia F, Dharamsi A, Howard OM, Cataisson C, Bliskovski V, Winston J, Feigenbaum L, Lichti U, Ruzicka T, Chavakis T, Yuspa SH (2010). "Gene from a psoriasis susceptibility locus primes the skin for inflammation". Sci Transl Med. 2 (61): 61ra90. doi:10.1126/scitranslmed.3001108. PMC 6334290. PMID 21148126.
- 1 2 Wolf R, Lewerenz V, Büchau AS, Walz M, Ruzicka T (2007). "Human S100A15 splice variants are differentially expressed in inflammatory skin diseases and regulated through Th1 cytokines and calcium". Exp. Dermatol. 16 (8): 685–91. doi:10.1111/j.1600-0625.2007.00587.x. PMID 17620096. S2CID 39343922.
- ↑ Hegyi Z, Zwicker S, Bureik D, Peric M, Koglin S, Batycka-Baran A, Prinz JC, Ruzicka T, Schauber J, Wolf R (2012). "Vitamin D analog calcipotriol suppresses the Th17 cytokine-induced proinflammatory S100 "alarmins" psoriasin (S100A7) and koebnerisin (S100A15) in psoriasis". J. Invest. Dermatol. 132 (5): 1416–24. doi:10.1038/jid.2011.486. PMID 22402441.
- ↑ Kulski JK, Lim CP, Dunn DS, Bellgard M (2003). "Genomic and phylogenetic analysis of the S100A7 (Psoriasin) gene duplications within the region of the S100 gene cluster on human chromosome 1q21". J. Mol. Evol. 56 (4): 397–406. Bibcode:2003JMolE..56..397K. doi:10.1007/s00239-002-2410-5. PMID 12664160. S2CID 39365917.
- 1 2 Wolf R, Voscopoulos CJ, FitzGerald PC, Goldsmith P, Cataisson C, Gunsior M, Walz M, Ruzicka T, Yuspa SH (2006). "The mouse S100A15 ortholog parallels genomic organization, structure, gene expression, and protein-processing pattern of the human S100A7/A15 subfamily during epidermal maturation". J. Invest. Dermatol. 126 (7): 1600–8. doi:10.1038/sj.jid.5700210. PMID 16528363.
- ↑ Murray JI, Tonkin ML, Whiting AL, Peng F, Farnell B, Cullen JT, Hof F, Boulanger MJ (2012). "Structural characterization of S100A7A reveals a novel zinc coordination site among S100 proteins and altered surface chemistry with functional implications for receptor binding". BMC Struct. Biol. 12: 16. doi:10.1186/1472-6807-12-16. PMC 3434032. PMID 22747601.
- ↑ Boeshans KM, Wolf R, Voscopoulos C, Gillette W, Esposito D, Mueser TC, Yuspa SH, Ahvazi B (2006). "Purification, crystallization and preliminary X-ray diffraction of human S100A15". Acta Crystallographica Section F. 62 (Pt 5): 467–70. doi:10.1107/S1744309106012838. PMC 2219979. PMID 16682778.
- ↑ "Entrez Gene: S100a7a S100 calcium binding protein".
- ↑ Briso EM, Guinea-Viniegra J, Bakiri L, Rogon Z, Petzelbauer P, Eils R, Wolf R, Rincón M, Angel P, Wagner EF (2013). "Inflammation-mediated skin tumorigenesis induced by epidermal c-Fos". Genes Dev. 27 (18): 1959–73. doi:10.1101/gad.223339.113. PMC 3792473. PMID 24029918.
- ↑ Nasser MW, Qamri Z, Deol YS, Ravi J, Powell CA, Trikha P, Schwendener RA, Bai XF, Shilo K, Zou X, Leone G, Wolf R, Yuspa SH, Ganju RK (2012). "S100A7 enhances mammary tumorigenesis through upregulation of inflammatory pathways". Cancer Res. 72 (3): 604–15. doi:10.1158/0008-5472.CAN-11-0669. PMC 3271140. PMID 22158945.
- ↑ Nasser MW, Wani N, Ahirwar DK, Powell CA, Ravi J, Elbaz MM, Zhao H, Padilla L, Zhang X, Shilo K, Ostrowski MC, Shapiro CL, Carson WE, Ganju RK (2015). "RAGE mediates S100A7-induced breast cancer growth and metastasis by modulating the tumor microenvironment". Cancer Res. 75 (6): 974–85. doi:10.1158/0008-5472.CAN-14-2161. PMC 4359968. PMID 25572331.
External links
- Overview of all the structural information available in the PDB for UniProt: Q86SG5 (Protein S100-A7A) at the PDBe-KB.
PDB gallery | |
|---|---|
|