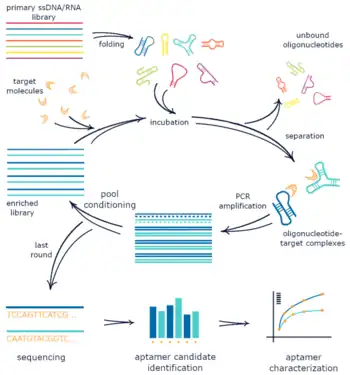
Systematic evolution of ligands by exponential enrichment (SELEX), also referred to as in vitro selection or in vitro evolution, is a combinatorial chemistry technique in molecular biology for producing oligonucleotides of either single-stranded DNA or RNA that specifically bind to a target ligand or ligands. These single-stranded DNA or RNA are commonly referred to as aptamers.[1][2][3] Although SELEX has emerged as the most commonly used name for the procedure, some researchers have referred to it as SAAB (selected and amplified binding site) and CASTing (cyclic amplification and selection of targets)[4][5] SELEX was first introduced in 1990. In 2015, a special issue was published in the Journal of Molecular Evolution in the honor of quarter century of the discovery of SELEX.[6]
The process begins with the synthesis of a very large oligonucleotide library, consisting of randomly generated sequences of fixed length flanked by constant 5' and 3' ends. The constant ends serve as primers, while a small number of random regions are expected to bind specifically to the chosen target. For a randomly generated region of length n, the number of possible sequences in the library using conventional DNA or RNA is 4n (n positions with four possibilities (A,T,C,G) at each position). The sequences in the library are exposed to the target ligand - which may be a protein or a small organic compound - and those that do not bind the target are removed, usually by affinity chromatography or target capture on paramagnetic beads.[7] The bound sequences are eluted and amplified by PCR[2][3] to prepare for subsequent rounds of selection in which the stringency of the elution conditions can be increased to identify the tightest-binding sequences.[2] A caution to consider in this method is that the selection of extremely high, sub-nanomolar binding affinity entities may not in fact improve specificity for the target molecule.[8] Off-target binding to related molecules could have significant clinical effects.
SELEX has been used to develop a number of aptamers that bind targets interesting for both clinical and research purposes.[9] Nucleotides with chemically modified sugars and bases have been incorporated into SELEX reactions to increase the chemical diversity at each base, expanding the possibilities for specific and sensitive binding, or increasing stability in serum or in vivo.[9][10]
Procedure
Aptamers have emerged as a novel category in the field of bioreceptors due to their wide applications ranging from biosensing to therapeutics. Several variations of their screening process, called SELEX have been reported which can yield sequences with desired properties needed for their final use.[11]
Generating single stranded oligonucleotide library
The first step of SELEX involves the synthesis of fully or partially randomized oligonucleotide sequences of some length flanked by defined regions which allow PCR amplification of those randomized regions and, in the case of RNA SELEX, in vitro transcription of the randomized sequence.[2][3][12] While Ellington and Szostak demonstrated that chemical synthesis is capable of generating ~1015 unique sequences for oligonucleotide libraries in their 1990 paper on in vitro selection,[3] they found that amplification of these synthesized oligonucleotides led to significant loss of pool diversity due to PCR bias and defects in synthesized fragments.[3] The oligonucleotide pool is amplified and a sufficient amount of the initial library is added to the reaction so that there are numerous copies of each individual sequence to minimize the loss of potential binding sequences due to stochastic events.[3] Before the library is introduced to target for incubation and selective retention, the sequence library must be converted to single stranded oligonucleotides to achieve structural conformations with target binding properties.[2][3]
Target incubation
Immediately prior to target introduction, the single stranded oligonucleotide library is often heated and cooled slowly to renature oligonucleotides into thermodynamically stable secondary and tertiary structures.[3][7] Once prepared, the randomized library is incubated with immobilized target to allow oligonucleotide-target binding. There are several considerations for this target incubation step, including the target immobilization method and strategies for subsequent unbound oligonucleotide separation, incubation time and temperature, incubation buffer conditions, and target versus oligonucleotide concentrations. Examples of target immobilization methods include affinity chromatography columns,[3] nitrocellulose binding assay filters,[2] and paramagnetic beads.[7] Recently, SELEX reactions have been developed where the target is whole cells, which are expanded near complete confluence and incubated with the oligonucleotide library on culture plates.[13] Incubation buffer conditions are altered based on the intended target and desired function of the selected aptamer. For example, in the case of negatively charged small molecules and proteins, high salt buffers are used for charge screening to allow nucleotides to approach the target and increase the chance of a specific binding event.[3] Alternatively, if the desired aptamer function is in vivo protein or whole cell binding for potential therapeutic or diagnostic application, incubation buffer conditions similar to in vivo plasma salt concentrations and homeostatic temperatures are more likely to generate aptamers that can bind in vivo. Another consideration in incubation buffer conditions is non-specific competitors. If there is a high likelihood of non-specific oligonucleotide retention in the reaction conditions, non specific competitors, which are small molecules or polymers other than the SELEX library that have similar physical properties to the library oligonucleotides, can be used to occupy these non-specific binding sites.[13] Varying the relative concentration of target and oligonucleotides can also affect properties of the selected aptamers. If a good binding affinity for the selected aptamer is not a concern, then an excess of target can be used to increase the probability that at least some sequences will bind during incubation and be retained. However, this provides no selective pressure for high binding affinity, which requires the oligonucleotide library to be in excess so that there is competition between unique sequences for available specific binding sites.[2]
Binding sequence elution and amplification
Once the oligonucleotide library has been incubated with target for sufficient time, unbound oligonucleotides are washed away from immobilized target, often using the incubation buffer so that specifically bound oligonucleotides are retained.[3] With unbound sequences washed away, the specifically bound sequences are then eluted by creating denaturing conditions that promote oligonucleotide unfolding or loss of binding conformation including flowing in deionized water,[3] using denaturing solutions containing urea and EDTA,[13][14] or by applying high heat and physical force.[7] Upon elution of bound sequences, the retained oligonucleotides are reverse-transcribed to DNA in the case of RNA or modified base selections,[2][3][13] or simply collected for amplification in the case of DNA SELEX.[15] These DNA templates from eluted sequences are then amplified via PCR and converted to single stranded DNA, RNA, or modified base oligonucleotides, which are used as the initial input for the next round of selection.[2][3]
Obtaining ssDNA
One of the most critical steps in the SELEX procedure is obtaining single stranded DNA (ssDNA) after the PCR amplification step. This will serve as input for the next cycle so it is of vital importance that all the DNA is single stranded and as little as possible is lost. Because of the relative simplicity, one of the most used methods is using biotinylated reverse primers in the amplification step, after which the complementary strands can be bound to a resin followed by elution of the other strand with lye. Another method is asymmetric PCR, where the amplification step is performed with an excess of forward primer and very little reverse primer, which leads to the production of more of the desired strand. A drawback of this method is that the product should be purified from double stranded DNA (dsDNA) and other left-over material from the PCR reaction. Enzymatic degradation of the unwanted strand can be performed by tagging this strand using a phosphate-probed primer, as it is recognized by enzymes such as Lambda exonuclease. These enzymes then selectively degrade the phosphate tagged strand leaving the complementary strand intact. All of these methods recover approximately 50 to 70% of the DNA. For a detailed comparison refer to the article by Svobodová et al. where these, and other, methods are experimentally compared.[16] In classical SELEX, the process of randomized single stranded library generation, target incubation, and binding sequence elution and amplification described above are repeated until the vast majority of the retained pool consists of target binding sequences,[2][3] though there are modifications and additions to the procedure that are often used, which are discussed below.
Negative or counter selection
In order to increase the specificity of aptamers selected by a given SELEX procedure, a negative selection, or counter selection, step can be added prior to or immediately following target incubation. To eliminate sequences with affinity for target immobilization matrix components from the pool, negative selection can be used where the library is incubated with target immobilization matrix components and unbound sequences are retained.[14][17][15] Negative selection can also be used to eliminate sequences that bind target-like molecules or cells by incubating the oligonucleotide library with small molecule target analogs, undesired cell types, or non-target proteins and retaining the unbound sequences.[13][15][18]
Tracking selection progression
To track the progress of a SELEX reaction, the number of target bound molecules, which is equivalent to the number of oligonucleotides eluted, can be compared to the estimated total input of oligonucleotides following elution at each round.[3][19] The number of eluted oligonucleotides can be estimated through elution concentration estimations via 260 nm wavelength absorbance[19] or fluorescent labeling of oligonucleotides.[7] As the SELEX reaction approaches completion, the fraction of the oligonucleotide library that binds target approaches 100%, such that the number of eluted molecules approaches the total oligonucleotide input estimate, but may converge at a lower number.[3]
Caveats and considerations
Some SELEX reactions can generate probes that are dependent on primer binding regions for secondary structure formation.[7] There are aptamer applications for which a short sequence, and thus primer truncation, is desirable.[20] An advancement on the original method allows an RNA library to omit the constant primer regions, which can be difficult to remove after the selection process because they stabilize secondary structures that are unstable when formed by the random region alone.[21]
Chemically modified nucleotides
Recently, SELEX has expanded to include the use of chemically modified nucleotides. These chemically modified oligonucleotides offer many potential advantages for selected aptamers including greater stability and nuclease resistance, enhanced binding for select targets, expanded physical properties - like increased hydrophobicity, and more diverse structural conformations.[9][10][22]
The genetic alphabet, and thus possible aptamers, is also expanded using unnatural base pairs[23][24] the use of these unnatural base pairs was applied to SELEX and high affinity DNA aptamers were generated.[25]
SELEX variants and alternative aptamer selection methods
FRELEX was developed in 2016 by NeoVentures Biotechnology Inc to allow the selection of aptamers without immobilizing the target or the oligonucleotide library.[26] Immobilization is a necessary component of SELEX; however, it has the potential to inhibit key epitopes, and thus weaken the likelihood of successful binding, particularly when working with small molecules.[27][28] FRELEX follows a similar overall methodology to SELEX; however, instead of immobilizing the target, the researcher introduces a series of random and blocker oligonucleotides to an immobilization field before introduction to the target.[26] This allows the researcher to better target small molecules that may be lost during partitioning.[26] It also can be used in some circumstances to select an aptamer library without knowing the target.[29]
Prior targets
The technique has been used to evolve aptamers of extremely high binding affinity to a variety of target ligands, including small molecules such as ATP[30] and adenosine[12][31] and proteins such as prions[32] and vascular endothelial growth factor (VEGF).[33] Moreover, SELEX has been used to select high-affinity aptamers for complex targets such as tumor cells,[34][35] tumor exosomes,[36][37] or tumor tissue.[38] Clinical uses of the technique are suggested by aptamers that bind tumor markers,[39] GFP-related fluorophores,[40] and a VEGF-binding aptamer trade-named Macugen has been approved by the FDA for treatment of macular degeneration.[33][41] Additionally, SELEX has been utilized to obtain highly specific catalytic DNA or DNAzymes. Several metal-specific DNAzymes have been reported including the GR-5 DNAzyme (lead-specific),[42] the CA1-3 DNAzymes (copper-specific),[43] the 39E DNAzyme (uranyl-specific) [44] and the NaA43 DNAzyme (sodium-specific).[45]
These developed aptamers have seen diverse application in therapies for macular degeneration[41] and various research applications including biosensors,[20] fluorescent labeling of proteins[46] and cells,[47] and selective enzyme inhibition.[48]
See also
- Aptamer – Oligonucleotide or peptide molecules that bind specific targets
- Deoxyribozyme – DNA oligonucleotides that can perform a specific chemical reaction
- Anti-thrombin aptamers – Oligonucleotides which recognize the exosites of human thrombin
- Bacterial one-hybrid system – Method for identifying the sequence-specific target site of a DNA-binding domain
References
- ↑ Hak-Hagir A (1978). "[The Hak-Hagir skin conduit]". Zeitschrift für Urologie und Nephrologie. 71 (9): 639–642. PMID 362762.
- 1 2 3 4 5 6 7 8 9 10 Tuerk C, Gold L (August 1990). "Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage T4 DNA polymerase". Science. 249 (4968): 505–10. Bibcode:1990Sci...249..505T. doi:10.1126/science.2200121. PMID 2200121.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 Ellington AD, Szostak JW (August 1990). "In vitro selection of RNA molecules that bind specific ligands". Nature. 346 (6287): 818–22. Bibcode:1990Natur.346..818E. doi:10.1038/346818a0. PMID 1697402. S2CID 4273647.
- ↑ Blackwell TK, Weintraub H (November 1990). "Differences and similarities in DNA-binding preferences of MyoD and E2A protein complexes revealed by binding site selection". Science. 250 (4984): 1104–10. Bibcode:1990Sci...250.1104B. doi:10.1126/science.2174572. PMID 2174572. S2CID 1995608.
- ↑ Wright WE, Binder M, Funk W (August 1991). "Cyclic amplification and selection of targets (CASTing) for the myogenin consensus binding site". Molecular and Cellular Biology. 11 (8): 4104–10. doi:10.1128/mcb.11.8.4104. PMC 361222. PMID 1649388.
- ↑ Gold L (December 2015). "SELEX: How It Happened and Where It will Go". Journal of Molecular Evolution. 81 (5–6): 140–143. Bibcode:2015JMolE..81..140G. doi:10.1007/s00239-015-9705-9. PMC 4661202. PMID 26480964.
- 1 2 3 4 5 6 Stoltenburg R, Schubert T, Strehlitz B (2015-07-29). "In vitro Selection and Interaction Studies of a DNA Aptamer Targeting Protein A". PLOS ONE. 10 (7): e0134403. Bibcode:2015PLoSO..1034403S. doi:10.1371/journal.pone.0134403. PMC 4519192. PMID 26221730.
- ↑ Carothers JM, Oestreich SC, Szostak JW (June 2006). "Aptamers selected for higher-affinity binding are not more specific for the target ligand". Journal of the American Chemical Society. 128 (24): 7929–37. doi:10.1021/ja060952q. PMC 4287982. PMID 16771507.
- 1 2 3 Wu YX, Kwon YJ (August 2016). "Aptamers: The "evolution" of SELEX". Methods. 106: 21–8. doi:10.1016/j.ymeth.2016.04.020. PMID 27109056.
- 1 2 Keefe AD, Cload ST (August 2008). "SELEX with modified nucleotides". Current Opinion in Chemical Biology. 12 (4): 448–56. doi:10.1016/j.cbpa.2008.06.028. PMID 18644461.
- ↑ Shorie M, Kaur H (October 2018). "Microtitre Plate Based Cell-SELEX Method". Bio-Protocol. 8 (20): e3051. doi:10.21769/BioProtoc.3051. PMC 8342047. PMID 34532522.
- 1 2 Huizenga DE, Szostak JW (January 1995). "A DNA aptamer that binds adenosine and ATP". Biochemistry. 34 (2): 656–65. doi:10.1021/bi00002a033. PMID 7819261.
- 1 2 3 4 5 Iwagawa T, Ohuchi SP, Watanabe S, Nakamura Y (January 2012). "Selection of RNA aptamers against mouse embryonic stem cells". Biochimie. 94 (1): 250–7. doi:10.1016/j.biochi.2011.10.017. PMID 22085640.
- 1 2 Vater A, Jarosch F, Buchner K, Klussmann S (November 2003). "Short bioactive Spiegelmers to migraine-associated calcitonin gene-related peptide rapidly identified by a novel approach: tailored-SELEX". Nucleic Acids Research. 31 (21): 130e–130. doi:10.1093/nar/gng130. PMC 275487. PMID 14576330.
- 1 2 3 Blank M, Weinschenk T, Priemer M, Schluesener H (May 2001). "Systematic evolution of a DNA aptamer binding to rat brain tumor microvessels. selective targeting of endothelial regulatory protein pigpen". The Journal of Biological Chemistry. 276 (19): 16464–8. doi:10.1074/jbc.M100347200. PMID 11279054. S2CID 39002909.
- ↑ Svobodová M, Pinto A, Nadal P, O' Sullivan CK (August 2012). "Comparison of different methods for generation of single-stranded DNA for SELEX processes". Analytical and Bioanalytical Chemistry. 404 (3): 835–42. doi:10.1007/s00216-012-6183-4. PMID 22733247. S2CID 206910212.
- ↑ Stoltenburg R, Reinemann C, Strehlitz B (October 2007). "SELEX--a (r)evolutionary method to generate high-affinity nucleic acid ligands". Biomolecular Engineering. 24 (4): 381–403. doi:10.1016/j.bioeng.2007.06.001. PMID 17627883.
- ↑ Haller AA, Sarnow P (August 1997). "In vitro selection of a 7-methyl-guanosine binding RNA that inhibits translation of capped mRNA molecules". Proceedings of the National Academy of Sciences of the United States of America. 94 (16): 8521–6. Bibcode:1997PNAS...94.8521H. doi:10.1073/pnas.94.16.8521. PMC 22984. PMID 9238009.
- 1 2 Sefah K, Shangguan D, Xiong X, O'Donoghue MB, Tan W (June 2010). "Development of DNA aptamers using Cell-SELEX". Nature Protocols. 5 (6): 1169–85. doi:10.1038/nprot.2010.66. PMID 20539292. S2CID 4953042.
- 1 2 Lubin AA, Hunt BV, White RJ, Plaxco KW (March 2009). "Effects of probe length, probe geometry, and redox-tag placement on the performance of the electrochemical E-DNA sensor". Analytical Chemistry. 81 (6): 2150–8. doi:10.1021/ac802317k. PMID 19215066.
- ↑ Jarosch F, Buchner K, Klussmann S (July 2006). "In vitro selection using a dual RNA library that allows primerless selection". Nucleic Acids Research. 34 (12): e86. doi:10.1093/nar/gkl463. PMC 1524915. PMID 16855281.
- ↑ Pinheiro VB, Taylor AI, Cozens C, Abramov M, Renders M, Zhang S, Chaput JC, Wengel J, Peak-Chew SY, McLaughlin SH, Herdewijn P, Holliger P (April 2012). "Synthetic genetic polymers capable of heredity and evolution". Science. 336 (6079): 341–4. Bibcode:2012Sci...336..341P. doi:10.1126/science.1217622. PMC 3362463. PMID 22517858.
- ↑ Kimoto M, Kawai R, Mitsui T, Yokoyama S, Hirao I (February 2009). "An unnatural base pair system for efficient PCR amplification and functionalization of DNA molecules". Nucleic Acids Research. 37 (2): e14. doi:10.1093/nar/gkn956. PMC 2632903. PMID 19073696.
- ↑ Yamashige R, Kimoto M, Takezawa Y, Sato A, Mitsui T, Yokoyama S, Hirao I (March 2012). "Highly specific unnatural base pair systems as a third base pair for PCR amplification". Nucleic Acids Research. 40 (6): 2793–806. doi:10.1093/nar/gkr1068. PMC 3315302. PMID 22121213.
- ↑ Kimoto M, Yamashige R, Matsunaga K, Yokoyama S, Hirao I (May 2013). "Generation of high-affinity DNA aptamers using an expanded genetic alphabet". Nature Biotechnology. 31 (5): 453–7. doi:10.1038/nbt.2556. PMID 23563318. S2CID 23329867.
- 1 2 3 10415034, Penner, Gregory & CA, "United States Patent: 10415034 - Method for the selection of aptamers for unbound targets", issued September 17, 2019
- ↑ Kohlberger M, Gadermaier G (August 2021). "SELEX: Critical factors and optimization strategies for successful aptamer selection". Biotechnology and Applied Biochemistry. 69 (5): 1771–1792. doi:10.1002/bab.2244. PMC 9788027. PMID 34427974. S2CID 237280042.
- ↑ Klapak D, Broadfoot S, Penner G, Singh A, Inapuri E (2018-10-11). "Development of novel aptamers for low-density lipoprotein particle quantification". PLOS ONE. 13 (10): e0205460. Bibcode:2018PLoSO..1305460K. doi:10.1371/journal.pone.0205460. PMC 6181373. PMID 30307996.
- ↑ Lecocq S, Spinella K, Dubois B, Lista S, Hampel H, Penner G (2018-01-05). de Franciscis V (ed.). "Aptamers as biomarkers for neurological disorders. Proof of concept in transgenic mice". PLOS ONE. 13 (1): e0190212. Bibcode:2018PLoSO..1390212L. doi:10.1371/journal.pone.0190212. PMC 5755763. PMID 29304088.
- ↑ Dieckmann T, Suzuki E, Nakamura GK, Feigon J (July 1996). "Solution structure of an ATP-binding RNA aptamer reveals a novel fold". RNA. 2 (7): 628–40. PMC 1369402. PMID 8756406.
- ↑ Burke DH, Gold L (May 1997). "RNA aptamers to the adenosine moiety of S-adenosyl methionine: structural inferences from variations on a theme and the reproducibility of SELEX". Nucleic Acids Research. 25 (10): 2020–4. doi:10.1093/nar/25.10.2020. PMC 146680. PMID 9115371.
- ↑ Mercey R, Lantier I, Maurel MC, Grosclaude J, Lantier F, Marc D (November 2006). "Fast, reversible interaction of prion protein with RNA aptamers containing specific sequence patterns". Archives of Virology. 151 (11): 2197–214. doi:10.1007/s00705-006-0790-3. PMID 16799875. S2CID 32195593.
- 1 2 Ulrich H, Trujillo CA, Nery AA, Alves JM, Majumder P, Resende RR, Martins AH (September 2006). "DNA and RNA aptamers: from tools for basic research towards therapeutic applications". Combinatorial Chemistry & High Throughput Screening. 9 (8): 619–32. doi:10.2174/138620706778249695. PMID 17017882.
- ↑ Daniels DA, Chen H, Hicke BJ, Swiderek KM, Gold L (December 2003). "A tenascin-C aptamer identified by tumor cell SELEX: systematic evolution of ligands by exponential enrichment". Proceedings of the National Academy of Sciences of the United States of America. 100 (26): 15416–21. Bibcode:2003PNAS..10015416D. doi:10.1073/pnas.2136683100. PMC 307582. PMID 14676325.
- ↑ Mayer G, Ahmed MS, Dolf A, Endl E, Knolle PA, Famulok M (December 2010). "Fluorescence-activated cell sorting for aptamer SELEX with cell mixtures". Nature Protocols. 5 (12): 1993–2004. doi:10.1038/nprot.2010.163. PMID 21127492. S2CID 4984082.
- ↑ Domenyuk V, Zhong Z, Stark A, Xiao N, O'Neill HA, Wei X, et al. (February 2017). "Plasma Exosome Profiling of Cancer Patients by a Next Generation Systems Biology Approach". Scientific Reports. 7 (1): 42741. Bibcode:2017NatSR...742741D. doi:10.1038/srep42741. PMC 5316983. PMID 28218293.
- ↑ Hornung T, O'Neill HA, Logie SC, Fowler KM, Duncan JE, Rosenow M, et al. (May 2020). "ADAPT identifies an ESCRT complex composition that discriminates VCaP from LNCaP prostate cancer cell exosomes". Nucleic Acids Research. 48 (8): 4013–4027. doi:10.1093/nar/gkaa034. PMC 7192620. PMID 31989173.
- ↑ Domenyuk V, Gatalica Z, Santhanam R, Wei X, Stark A, Kennedy P, et al. (March 2018). "Poly-ligand profiling differentiates trastuzumab-treated breast cancer patients according to their outcomes". Nature Communications. 9 (1): 1219. Bibcode:2018NatCo...9.1219D. doi:10.1038/s41467-018-03631-z. PMC 5865185. PMID 29572535.
- ↑ Ferreira CS, Matthews CS, Missailidis S (2006). "DNA aptamers that bind to MUC1 tumour marker: design and characterization of MUC1-binding single-stranded DNA aptamers". Tumour Biology. 27 (6): 289–301. doi:10.1159/000096085. PMID 17033199. S2CID 41664944.
- ↑ Paige JS, Wu KY, Jaffrey SR (July 2011). "RNA mimics of green fluorescent protein". Science. 333 (6042): 642–6. Bibcode:2011Sci...333..642P. doi:10.1126/science.1207339. PMC 3314379. PMID 21798953.
- 1 2 Vavvas D, D'Amico DJ (September 2006). "Pegaptanib (Macugen): treating neovascular age-related macular degeneration and current role in clinical practice". Ophthalmology Clinics of North America. 19 (3): 353–60. doi:10.1016/j.ohc.2006.05.008 (inactive 1 August 2023). PMID 16935210.
{{cite journal}}: CS1 maint: DOI inactive as of August 2023 (link) - ↑ Breaker RR, Joyce GF (December 1994). "A DNA enzyme that cleaves RNA". Chemistry & Biology. 1 (4): 223–9. doi:10.1016/1074-5521(94)90014-0. PMID 9383394.
- ↑ Carmi N, Shultz LA, Breaker RR (December 1996). "In vitro selection of self-cleaving DNAs". Chemistry & Biology. 3 (12): 1039–46. doi:10.1016/s1074-5521(96)90170-2. PMID 9000012.
- ↑ Liu J, Brown AK, Meng X, Cropek DM, Istok JD, Watson DB, Lu Y (February 2007). "A catalytic beacon sensor for uranium with parts-per-trillion sensitivity and millionfold selectivity". Proceedings of the National Academy of Sciences of the United States of America. 104 (7): 2056–61. Bibcode:2007PNAS..104.2056L. doi:10.1073/pnas.0607875104. PMC 1892917. PMID 17284609.
- ↑ Torabi SF, Wu P, McGhee CE, Chen L, Hwang K, Zheng N, Cheng J, Lu Y (May 2015). "In vitro selection of a sodium-specific DNAzyme and its application in intracellular sensing". Proceedings of the National Academy of Sciences of the United States of America. 112 (19): 5903–8. Bibcode:2015PNAS..112.5903T. doi:10.1073/pnas.1420361112. PMC 4434688. PMID 25918425.
- ↑ Umrao S, Jain V, Chakraborty B, Roy R (August 2018). "Protein-induced fluorescence enhancement as aptamer sensing mechanism for thrombin detection". Sensors and Actuators B: Chemical. 267: 294–301. doi:10.1016/j.snb.2018.04.039. S2CID 103202899.
- ↑ Terazono H, Anzai Y, Soloviev M, Yasuda K (April 2010). "Labelling of live cells using fluorescent aptamers: binding reversal with DNA nucleases". Journal of Nanobiotechnology. 8 (1): 8. doi:10.1186/1477-3155-8-8. PMC 2861636. PMID 20388214.
- ↑ Mondragón E, Maher LJ (September 2015). "RNA aptamer inhibitors of a restriction endonuclease". Nucleic Acids Research. 43 (15): 7544–55. doi:10.1093/nar/gkv702. PMC 4551934. PMID 26184872.
Further reading
- Levine HA, Nilsen-Hamilton M (February 2007). "A mathematical analysis of SELEX". Computational Biology and Chemistry. 31 (1): 11–35. doi:10.1016/j.compbiolchem.2006.10.002. PMC 2374838. PMID 17218151.
- Spill F, Weinstein ZB, Irani Shemirani A, Ho N, Desai D, Zaman MH (October 2016). "Controlling uncertainty in aptamer selection". Proceedings of the National Academy of Sciences of the United States of America. 113 (43): 12076–12081. arXiv:1612.08995. Bibcode:2016PNAS..11312076S. doi:10.1073/pnas.1605086113. PMC 5087011. PMID 27790993.