一氧化碳脱氢酶
一氧化碳脱氢酶(英語:,缩写为CODH)是一种存在于许多需氧和厌氧微生物中的一类脱氢酶,其催化的是一氧化碳(CO)氧化为二氧化碳(CO2)的反应或其逆反应,在这些微生物的代谢途径中扮演着关键的角色。[1]
- CO + H2O ⇌ CO2 + 2 H+ + 2 e−
| carbon-monoxide dehydrogenase (acceptor) | |||||||
|---|---|---|---|---|---|---|---|
| |||||||
| 识别码 | |||||||
| EC編號 | 1.2.99.2 | ||||||
| CAS号 | 64972-88-9 | ||||||
| 数据库 | |||||||
| IntEnz | IntEnz浏览 | ||||||
| BRENDA | BRENDA入口 | ||||||
| ExPASy | NiceZyme浏览 | ||||||
| KEGG | KEGG入口 | ||||||
| MetaCyc | 代谢路径 | ||||||
| PRIAM | 概述 | ||||||
| PDB | RCSB PDB PDBj PDBe PDBsum | ||||||
| 基因本体 | AmiGO / EGO | ||||||
| |||||||
不同来源的一氧化碳脱氢酶具有不同的结构和催化机理。需氧细菌中的一氧化碳脱氢酶是一类含钼(Mo)金属蛋白,需氧细菌利用它们在呼吸作用中氧化一氧化碳来获得能量。而在厌氧微生物,包括厌氧细菌和古菌中,其属于含镍(Ni)金属蛋白,在不同种属的微生物中发挥着不同的作用。特别在厌氧微生物固定一氧化碳和二氧化碳的Wood-Ljungdahl代谢途径中,一氧化碳脱氢酶可以与乙酰辅酶A合成酶(同为镍蛋白)结合形成双功能酶,共同发挥作用,将一氧化碳和二氧化碳转化为重要的代谢中间物乙酰辅酶A。[2]
由于一氧化碳脱氢酶能够高效地催化对动物和人体有很高毒性的一氧化碳氧化为二氧化碳,因此对于其催化机理的研究有助于治理环境中的一氧化碳污染。
分类
一氧化碳脱氢酶可以被分为两大类:存在于需氧微生物中的Mo-Fe-黄素(flavin)酶类和存在于厌氧微生物中的Ni-Fe酶类。由于在这两类金属酶中,位于活性位点起关键作用的金属分别是钼(Mo)和镍(Ni),因此分别称这两类酶为Mo-CODH和Ni-CODH。Mo-CODH对CO有很高的亲和力,[2] 适合于除去环境中痕量的一氧化碳气体;而就催化活性而言,Ni-CODH则是Mo-CODH的近1000倍。[3]
Mo-CODH
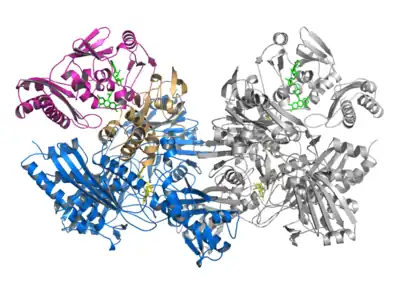
Mo-CODH存在于一类被称为一氧化碳自养菌(carboxydotrophs)的化能自养微生物中。此类细菌中的CODH是一类含Mo的铁硫黄素蛋白。与Ni-CODH相比,Mo-CODH对氧气不敏感。[2] 在嗜温菌Oligotropha carboxidovorans和嗜热菌Oligotropha thermocarboxydovorans中,Mo-CODH具有相似的分子量(230 kDa – 310 kDa)和相同的亚基组成。[4]
在一氧化碳自养菌中,除少量定位于细胞质外,大多数的Mo-CODH连接在细胞质膜的内表面上,[5] 从而有利于其与同样连在内膜上的其生理条件下的电子受体——细胞色素b561之间进行电子传递。[1] 在Mo-CODH中,Mo结合在molybdopterin胞嘧啶二核苷酸(MCD)辅因子上。序列分析显示Mo-CODH属于含Mo的羟化酶(hydroxylase)家族。[6] 以下以研究较多的O. carboxidovorans中的Mo-CODH为例进行详述。
亚基组成
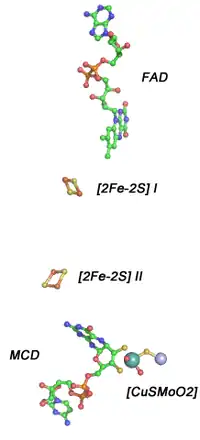
Mo-CODH由L(88.7 kDa,809个氨基酸)、M(30.2 kDa,288个氨基酸)、S(17.8 kDa,166个氨基酸)三个亚基组成。这三个亚基结合在一起形成“LMS复合物”,且LMS复合物两两结合形成(LMS)2,即Mo-CODH的天然活性形式为二聚体。[7] L亚基又被称为molybdoprotein,含有Mo和与之结合的MCD分子,其中Mo离子与另一金属离子Cu形成活性中心的[CuSMoO2]簇;M亚基为黄素蛋白(flavoprotein),其结合有黄素腺嘌呤二核苷酸(FAD);S亚基属于铁硫蛋白,结合了两个[2Fe-2S]簇。[7][8]
含有两个[2Fe-2S]簇的S亚基从结构上可以被分为N端和C端两个结构域,这两个结构域各结合了一个[2Fe-2S]簇。两个铁硫簇之间的距离约为12Å,在可以进行电子传递的最长距离(14 Å)之内。结合在C端结构域,靠近活性中心的铁硫簇被称为铁硫簇I([2Fe-2S] I),其被包埋在蛋白内部。结合在N端结构域远离活性中心的铁硫簇被称为铁硫簇II([2Fe-2S] II)。铁硫簇II位于S亚基和M亚基结合面上,暴露在溶剂环境中,距离FAD分子的异咯嗪环(isoalloxazine ring)大约9 Å,因此其功能被认为是在铁硫簇I和FAD分子之间传递电子。[7] 这两个结构域的折叠类型和结合铁硫簇II的氨基酸序列普遍存在于其他含Mo的羟化酶家族成员中。[9][10]
M亚基由N端、中间和C端三个结构域组成。其FAD分子的结合位点为两段特征性的“双甘氨酸(double-glycine)”序列,在含Mo的羟化酶家族中非常保守。[7] 其中一个双甘氨酸序列位于N端结构域的一段loop上,与FAD分子上的焦磷酸根相互作用。另一个双甘氨酸序列则位于中间结构域的一个小段α螺旋上与FAD分子相互作用。FAD分子结合在N端和中间结构域之间的缝隙中,且FAD的结合需要通过M亚基与L和S亚基作用所产生的构象变化来实现,游离的M亚基没有FAD的结合活性。[7][11] M亚基作为铁硫簇II与细胞色素b561之间电子传递的中间体,一方面接受来自铁硫簇II的电子,另一方面向细胞色素b561提供电子。[4]
分子量最大的L亚基不仅包含有活性位点,也在LMS复合物形成及Mo-CODH二体化中起主要作用。L亚基含有N端结构域和C端结构域,而Mo原子就定位在这两个结构域的结合面上。MCD辅因子的一端连接Mo的部分与活性中心的[CuSMoO2]簇一起结合在N端结构域的凹槽中;而辅因子的另一端则插入C端结构域的裂缝中。L亚基的N端结构域与M和S亚基相互作用以形成LMS复合物;而C端结构域则主要参与形成Mo-CODH二体结构。[7]
活性位点
Mo-CODH的活性位点位于L亚基,并被包埋于蛋白质内部,但从外部可以通过一个约17Å长的通道到达活性位点。这一通道由疏水残基构成,从而允许CO分子进入而防止极性溶剂分子(如水分子)进入。最初的研究认为活性位点的Mo与三个氧原子形成配位,且存在硒代巯基(SeH)共价连接在氨基酸残基Cys388上。[7] 然而随后的研究表明,Mo-CODH的活性位点是一个Cu-Mo金属簇,其包含有一个Cu离子和一个[MoO2],Cu离子和Mo离子之间通过一个配位硫原子(μS,又称为硫桥sulfido)来连接,结合在Cys388的Sγ原子上将此金属簇[CuSMoO2]与L亚基共价连接的是Cu而不是Se。[3][12] 而MCD是通过其上的两个S原子与Mo离子形成配位连接。这种由Mo和Cu两种金属组成的双核金属簇非常独特,因为所有已知的含molybdopterin或MCD的Mo金属蛋白中都是单核的。
无论Mo-CODH处在还原态还是氧化态,活性位点的Cu离子总是保持+I价,而连接Mo离子的MCD分子的构象也没有发生明显的变化。在空气中,Mo-CODH处于氧化态,此时Mo离子为+VI价,位于Mo离子与MCD的两个配位硫原子所形成平面垂直方向上的氧配基Oa与Mo离子之间形成双键连接;而与Mo离子配位的另一个氧配基Ob则有可能与之形成单键或双键。[3][12] 与氧化态的结构相比,还原态中Cu-Mo距离和Mo-Ob距离都有所延长。结合CO时,活性位点很可能是处于非常短暂的中间态,难以捕捉,因此研究者利用异腈基团与活性位点结合的结构来模拟中间态。由研究结果推测CO应该连接在Cu离子上而不直接与Mo连接。[12]
在Mo-CODH二聚体中,由于两个活性位点的间距超过了可以直接进行电子传递的距离,而且没有辅因子位于它们之间,两者之间不可能进行电子传递;因此Mo-CODH二聚体中的两个单体很可能可以独立进行催化活动,即每个单体都是一个完整的催化单位。[7]
Ni-CODH
Ni-CODH广泛存在于以一氧化碳为碳源和能源的厌氧微生物中,如乙酸生成细菌(acetogenic bacteria)、光养细菌(phototrophic bacteria)、氢气生成细菌(hydrogenogenic bacteria)、硫酸还原细菌和古菌(sulfate-reducing bacteria and archaea)以及甲烷生成古菌(methangenic archaea)。[1][13]
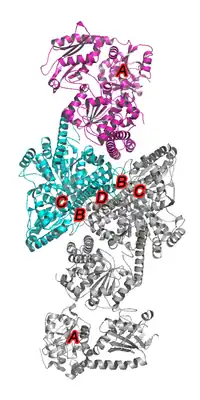
与Mo-CODH相比,Ni-CODH在活性位点处的关键金属为Ni,负责电子传递的铁硫簇为[4Fe-4S]而非[2Fe-2S],且没有发现有像Mo-CODH中的FAD和MCD辅因子,整体结构上也没有相似性。除此之外,不同来源的Ni-CODH差别很大:既有单功能的一氧化碳脱氢酶只负责催化CO的氧化;又有双功能酶,即同时具有一氧化碳脱氢酶和乙酰辅酶A合成酶(ACS)的活性,生理状态下需要Ni-CODH与ACS结合共同发挥功能。因此,又常常以CODH/ACS复合物的名称来代替Ni-CODH。与Mo-CODH相似的是,CODH/ACS也需要形成二聚体来发挥催化作用。
分类
基于催化活性、代谢功能和蛋白性质及组成,可将不同的CODH/ACS分为四类:[14][15]
- 第一类是存在于专性化能自养的甲烷生成古菌的CODH/ACS,负责直接利用CO2和氢气(H2)来合成乙酰辅酶A;
- 第二类酶则负责与第一类相反的反应,即将乙酰辅酶A分解为CO2和甲烷(CH4),其主要存在于利用乙酸的甲烷生成古菌和硫酸还原菌中;
- 第三类CODH/ACS发现存在于乙酸生成细菌中,其作用与第一类酶相同,只是以丙酮酸盐作为CO2和H2的来源,并参与Wood-Ljungdahl途径;
- 第四类酶是一类单功能酶,只负责催化CO的氧化。
虽然把CODH/ACS家族分为这样四类与利用生物信息学进行的序列分析的结果相符合;[15] 但对来自M. thermophila的CODH/ACS(第二类酶)进行研究发现,乙酰辅酶A的合成和分解都是由相同的CODH/ACS来完成,[16] 这说明这一分类的依据和方法还需进一步改进。
亚基组成
这四类酶的亚基组成也有较大差别。[14] 第一/二类CODH/ACS都由五个不同的亚基(αβγδε)组成。第三类CODH/ACS则由α和β两个亚基组成,其中α亚基与第一/二类中的β亚基同源,但α亚基多了N端30 kDa的一段序列;而其β亚基与第一/二类中的α亚基同源,但同源性较差,且相比之下,β亚基的分子量要小一些。虽然第三类的来源菌中也存在与第一/二类CODH/ACS对应的γ和δ亚基,但不存在ε亚基的同源蛋白,且γ和δ亚基不参与CODH/ACS的组成,而是形成异二聚体发挥甲基传递作用。第四类CODH/ACS中只含有与第三类中β亚基具有很高同源性的单个蛋白。
活性位点
由于除了第四类CODH/ACS外,其他类酶都是双功能,甚至多功能酶,从而具有两个或多个活性位点。其中,两个核心反应,即CO的氧化和乙酰辅酶A的合成,对应的活性位点金属簇分别为C簇[17] 和A簇。其中,C簇位于α亚基(第三类CODH/ACS中为β亚基),其组成为[Ni-4Fe-4S]或[Ni-4Fe-5S];[18] 而A簇位于β亚基(第三类CODH/ACS中为α亚基)。A簇和C簇之间通过一个疏水通道相连,从而可以相互传递催化产物。[19][20] 此外,α亚基(第三类CODH/ACS中为β亚基)二聚体中还含有5-7个铁硫簇[4Fe-4S],负责电子传递。[14]
催化机理
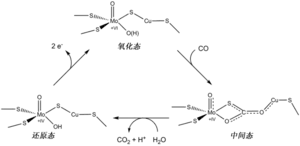
负责CO的氧化的活性位点C簇的催化机制主要由两步反应组成。[2] 第一步,CO被氧化,而C簇被还原;第二步,电子从C簇传递到蛋白外的电子受体,如ferredoxin蛋白,C簇回到初始状态。在这一过程,C簇中存在四种氧化还原态:Cox,Cred1,Cint和Cred2。其中,处于氧化状态(Ni2+-Fe13+)的Cox在获得一个电子之后转化为Cred1。Cred1被证明是与CO和OH基团结合时C簇的状态;而Cred2出现在Cred1之后,此时CO可能已被氧化为CO2,并释放出H+离子。这步反应可能是由OH基团攻击连接在Ni上的CO引发,因为在C簇周围有多个碱性残基(包括一个赖氨酸和多个组氨酸),使结合在Fe1上的水分子的pKa值显著降低,从而使水分子转变为活性的OH基团,对CO进行攻击。此外,在C簇附近还有一个保守的天冬氨酸,可以与组氨酸形成氢键连接,促进此酸碱反应的发生。反应生成的H+离子通过组氨酸形成的质子通道传递到外部溶剂中。CO被氧化为CO2后,C簇接受两个电子从Cred1转变为Cred2,随后两个电子一个接一个地被传递到B簇。传走第一个电子后,C簇转化为中间态Cint;两个电子都传到B簇后,C簇又重新转化为Cred1,从而完成一次催化循环。
參考資料
- (英文)Ferry, J.G. . Annu Rev Microbiol. 1995, 49: 305–333.
- (英文)Ragsdale, S.W. . Crit Rev Biochem Mol Biol. 2004, 39: 165–195.
- (英文)Gnida, M.; et al. . Biochemistry. 2003, 42: 222–230.
- (英文)Meyer, O., S. Jacobitz, and B. Kruger. . FEMS Microbiol Rev. 1986, 39: 161–179.
- (英文)Rohde, M., F. Mayer, and O. Meyer. . J Biol Chem. 1984, 259: 14788–14792.
- (英文)Schubel, U.; et al. . J Bacteriol. 1995, 177: 2197–2203.
- (英文)Dobbek, H.; et al. . Proc Natl Acad Sci U S A. 1999, 96: 8884–8889.
- (英文)Meyer, O.; et al. . Biol Chem. 2000, 381: 865–876.
- (英文)Romao, MJ; et al. . Science. 1995, 270: 1170–1176.
- (英文)Rebelo, J; et al. . J Mol Biol. 2000, 297: 135–146.
- (英文)Gremer, L; et al. . J Biol Chem. 2000, 275: 1864–1872.
- (英文)Dobbek, H.; et al. . Proc Natl Acad Sci U S A. 2002, 99: 15971–15976.
- (英文)Ragsdale, S.W. and Kumar, M. . Chem Rev. 1996, 96: 2515–2540.
- (英文)Lindahl, P.A. . Biochemistry. 2002, 41: 2097–2105.
- (英文)Lindahl, P.A. and B. Chang. . Orig Life Evol Biosph. 2001, 31: 403–434.
- (英文)Grahame, D.A., S. Gencic, and E. DeMoll. . Arch Microbiol. 2005, 184: 32–40.
- (英文)Kumar, M.; et al. . Journal of the American Chemical Society. 1993, 115: 11646–11647.
- (英文)Dobbek, H.; et al. . J Am Chem Soc. 2004, 126: 5382–5387.
- (英文)Doukov, T.I.; et al. . Science. 2002, 298: 567–572.
- (英文)Darnault, C.; et al. . Nat Struct Biol. 2003, 10: 271–279.