共生體學說
共生体學說(英語:),又稱內共生學說(英語:),是關於真核生物細胞中的一些自主胞器ㄧ粒線體和葉綠體起源的學説。根據這個學説,它們起源於共生於真核生物細胞中(之内)的原核生物。這種理論認爲粒線體起源於好氧細菌(很可能是接近於立克次體的變形菌門細菌,特別是远洋杆菌目()[1][2]),而葉綠體源於内共生的光合自營原核生物的藍細菌。這個理論的證據非常完整,目前已經被廣泛接受。
歷史
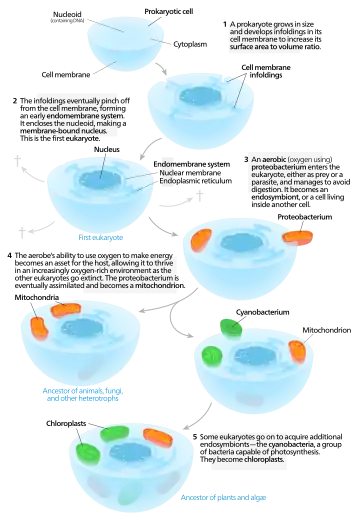
線粒體和質體起源的一個模型。
1905年,康斯坦丁·梅列施柯夫斯基最先提出葉綠體是由原先的内共生體形成的這一想法,隨後1920年代伊凡·沃林提出了對線粒體的相同想法。隨着人們發現它們含有DNA,這些想法被Henry Ris重新提出。
内共生假説被琳·馬古利斯所普及。在她1981年的《細胞進化中的共生》中,她認爲真核細胞起源於相互作用的個體組成的群落,也包括内共生的螺旋體形成了真核生物的鞭毛和纖毛。后一種想法沒有被廣泛接受,因爲鞭毛缺乏DNA,在結構之外和原核生物也沒有明顯相似性。根據馬古利斯和多里昂·薩根(1996),“生命並不是通過戰鬥,而是通過協作佔據整個全球的”,而達爾文關於進化由競爭驅動的想法是不完善的。然而也有人認爲内共生更像奴隸制而非互利共生。
有人認爲過氧化物酶體可能也具有内共生起源,儘管它們缺乏DNA。克里斯汀·德·迪夫認爲它們也許是最早的内共生體,使得細胞能夠抵抗地球大氣中越來越多的氧氣。然而,現在看來它們可能是“從頭合成”的,而非具有共生起源。
證據
線粒體和葉綠體起源于古代細菌内共生的證據如下:[3][4][5][6]
- 它們被兩層或更多的膜所包被,其中最裏面一層的成分與細胞中其它膜的都不同,而更接近於原核生物的細胞膜。另外有更多層的多次內共生生物的存在被發現。
- 新的線粒體和葉綠體只能通過類似二分分裂的過程自己形成。在一些藻類以及眼蟲(Euglena)中,可以用藥物或長時間缺乏光照來破壞葉綠體而同時不影響細胞。這種情況下,細胞喪失的葉綠體將不能夠自行再生。
- 細胞器的核糖體和細菌相似,細菌的核糖體是70S,線粒體的核糖體是55S。
- 葉綠體的很多内部結構和生物化學特徵,如類囊体的存在和某些葉綠素和藍細菌使用的蛋白質很接近。
- 可以抑制真细菌蛋白质合成的抗生素也可以抑制线粒体和叶绿体蛋白质的合成,但不能抑制真核生物细胞质的蛋白质合成。
- 對細菌、葉綠體和真核生物基因組構件的系統發生樹同樣支持了葉綠體與藍細菌更接近。
- 葉綠體存在於很多完全不同的原生生物中,這些生物普遍和不包含葉綠體的原生生物更接近。這表明了,如果葉綠體起源於細胞的一個部分,很難解釋他們多次分開起源而互相又非常接近的假設。
- 一些核中編碼的蛋白被轉運到細胞器中,而線粒體和葉綠體的基因組相對於其它生物來說都小得多。這和内共生物形成後越來越依賴真核生物宿主相一致。
- DNA序列分析和系統發生學表明了核DNA包含了一些可能來源於外部的基因片段,極可能是葉綠體的基因。
- 線粒體和葉綠體都含有DNA,這些DNA與細胞核中的很不同,卻類似細菌的DNA(共价、闭合、环状的形状及其大小)。
- 線粒體具有和真核宿主細胞不同的遺傳密碼,這些密碼與細菌和古菌中的使用的編碼很類似。
- 細胞器的大小與細菌相當。
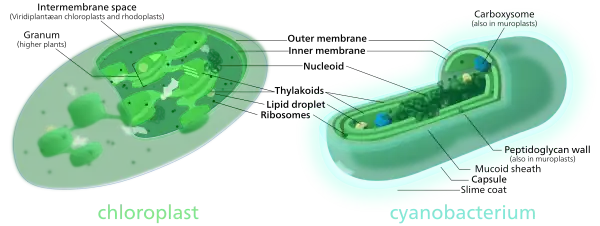
對比比較葉綠體(左)和藍細菌(右)顯示其相似之處。
多重内共生
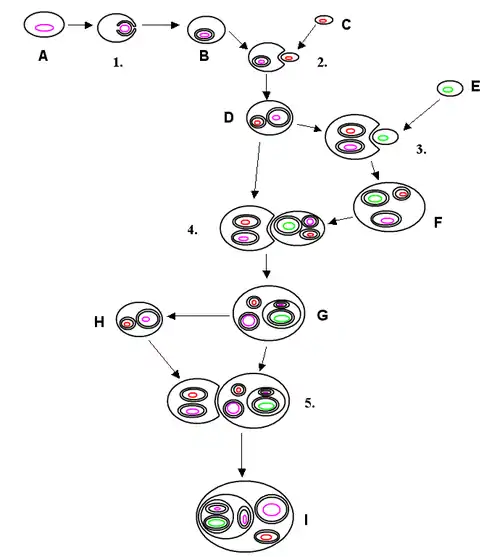 |
圖中顔色
|
吞噬紅藻所進行的多次內共生較為複雜,其分子生物學證據分歧較大,暫不清楚這一過程獨立發生了多少次。在隱藻的質體中通常會出現核型體這一遺跡結構,其作用為表達SLEMA相關蛋白質。雙鞭毛蟲的質體只有三層膜結構。
參看
- 地衣
- 原始生命體
- 連續內共生學說
參考文獻
- Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts and Peter Walter, Molecular Biology of the Cell, Garland Science, New York, 2002. ISBN 0-8153-3218-1. (General textbook)
- Jeffrey L. Blanchard and Michael Lynch (2000), "Organellar genes: why do they end up in the nucleus?", Trends in Genetics, 16 (7), pp. 315-320. (Discusses theories on how mitochondria and chloroplast genes are transferred into the nucleus, and also what steps a gene needs to go through in order to complete this process.)
- Paul Jarvis (2001), "Intracellular signalling: The chloroplast talks!", Current Biology, 11 (8), pp. R307-R310. (Recounts evidence that chloroplast-encoded proteins affect transcription of nuclear genes, as opposed to the more well-documented cases of nuclear-encoded proteins that affect mitochondria or chloroplasts.)
- Zimorski V, Ku C, Martin WF, Gould SB. 2014. Endosymbiotic theory for organelle origins. Current Opinion in Microbiology 22:33-48.https://pubmed.ncbi.nlm.nih.gov/25306530/
- . ScienceDaily. July 25, 2011 [2011-07-26]. (原始内容存档于2020-11-14).
- J. Cameron Thrash; et al. . Scientific Reports. 2011. doi:10.1038/srep00013.
- (页面存档备份,存于) Kimball, J. 2010. Kimball's Biology Pages. Accessed October 13, 2010. An online open source biology text by Harvard professor, and author of a general biology text, John W. Kimball.
- Reece, J., Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson, 2010. Campbell Biology. 9th Edition Benjamin Cummings; 9th Ed. (October 7, 2010)
- Raven, P., George Johnson, Kenneth Mason, Jonathan Losos, Susan Singer, 2010. Biology. McGraw-Hill 9th Ed. (January 14, 2010)
- 吴相钰; 陈守良; 葛明德. 4. 高等教育出版社. 2014. ISBN 9787040396317.
This article is issued from Wikipedia. The text is licensed under Creative Commons - Attribution - Sharealike. Additional terms may apply for the media files.