藻類
藻類(Algae)是一非正式术语,指的是大量多样的光合真核生物复系群,包括来自多个不同演化枝(支序群)的物种。它們是能行光合作用的生物,並且一些藻類與比較高等的植物有關。雖然其他藻類看似從藍綠菌得到光合作用的能力,但是在演化上有獨立的分支。所有藻類缺乏真的根、莖、葉和其他可在高等植物上發現的組織構造。藻類與細菌和原生動物不同之處,是藻類產生能量的方式為光合自營。
多種光合真核生物的非正式術語 化石时期: | |
|---|---|
 | |
| 許多不同的藻類,生長環境從淺水到海床 | |
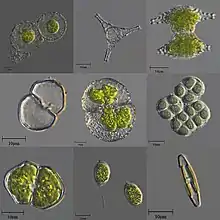 | |
| 各種微觀的單細胞和群體淡水藻類 | |
| 科学分类 | |
| Missing taxonomy template(修正): | 真核生物 |
| 传统分类中包含的子类群 | |
| 一般會排除 | |
| |




藻類本身並不獨立為一生物“界”,而是分散於真核生物域中各界。屬於原生生物界的藻類有裸藻、甲藻(或稱渦鞭毛藻)、隱藻、金黃藻(包括矽藻等浮游藻)、紅藻、綠藻和褐藻。而生殖構造複雜的輪藻則屬於植物界。屬於大型藻者一般僅有紅藻、綠藻和褐藻等為大型肉眼可顯而易見之固著性藻類。此類大型藻幾乎99%以上之種類棲息於海水環境中,故大型藻多以海藻稱之。另外,有些肉眼可見的固著性藍綠藻和少數之矽藻嚴格而言應該亦屬於大型藻的範圍。
各種藻類的差異很大,人類許多傳統應用以及工業應用都會用到藻類。傳統的藻類養殖已進行了上千年,也是東亞飲食文化中的傳統。近來除了可食用藻類之外,藻類還有其他的農藝應用,例如作牲畜的飼料、用藻類來作生物修復或污染控制、將太陽能轉換為藻類生質燃料或是其他工業上會用到的化學品,以及醫學或是科技上的應用。2020的回顧報告發現,藻類可以在碳截存(carbon sequestration)上有重要的作用,以緩解氣候變化,同時也為全球的經濟體提供加值的商品[3]。
生態
藻類在水裡非常常見,在陸域環境也是。然而陸域藻類通常較不顯眼,且於潮濕、熱帶地區比乾燥地區更常見,因為藻類缺乏維管束和其他營陸地生活的適應構造。藻類在其他地點如雪地或以地衣的形式在裸露岩石表面與真菌共生。
種類繁複的藻類在水域生態系扮演重要角色。微觀下懸浮於水柱者﹝浮游植物﹞提供食物給大多數海洋食物鏈。當藻類密度非常高﹝水華﹞時,可能使水變色,與其他生物競爭或使其他生物中毒或窒息。海草大部分生長在淺海水中,然而有些已有生長於300公尺深的紀錄。[4]有些供人類食用或生產有用物質如洋菜、鹿角菜膠或肥料。
分類
雖然傳統上藍綠藻歸於藻類,稱為藍綠藻門(Cyanophyte),近年研究通常將它排除,因為和藻類差異甚大,諸如缺乏膜包圍的胞器,含有單一環狀染色體,細胞壁含有肽聚糖,核糖體大小、成分和真核生物不同。[6][7]藍綠藻在特化摺疊的原生質膜(稱為葉綠囊膜)上行光合作用。因此,它們即使擁有類似的生態區位,彼此仍然差異很大所以現時被歸屬細菌界中。
William Henry Harvey(1811—1866) 首先根據色素將藻類分為四類,首次將生化準則用於植物分類,其四類為:紅藻、褐藻、綠藻和矽藻。[8]
依現在的定義,藻類是真核生物,在稱為葉綠體的膜狀胞器內行光合作用。葉綠體內含環狀DNA,結構和藍綠藻相似,可能代表退化的藍綠藻內共生胞器。各演化系(lineage)藻類葉綠體的確切特性均不相同,反映出不同的內共生事件。下表列出三個主要群組,它們的演化系關係表示於左側。需注意許多群組均包含不再行光合作用的成員。有些仍保有色素體但非葉綠體,有些則已經完全喪失。
種系發生:[9]
|
藍藻 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 主要群組 | 成員 | 內共生體 | 说明 |
|---|---|---|---|
| 广义植物/ 原始色素体生物 |
藍綠藻 | 含有初級葉綠體,i.e,葉綠體由雙層膜包圍,可能源自單一內共生事件。紅藻的葉綠體含葉綠素a和d(通常),和藻胆素,而綠藻的含葉綠素a和b。高等植物的色素近似綠藻,可能由綠藻演化而來。所以綠藻門是植物界的親近分類,有時會歸類為綠植界(Virdiplantae)。 | |
| 古虫界与有孔蟲界 |
|
綠藻 | 擁有內含葉綠素a和b的綠色葉綠體[6]。葉綠體被3或4層膜包圍,可能分別源自吞入的綠藻。
绿蛛网藻,屬於絲足蟲門。包含小型的類核體,其為藻類細胞核的殘餘。 裸藻綱,屬於眼虫門。主要生活於淡水,葉綠體有3層膜。有人認為其內共生綠藻是經由myzocytosis而非吞噬作用(phagocytosis) |
| 色藻界和囊泡虫类 |
|
紅藻 | 擁有內含葉綠素a和c、藻胆素的葉綠體。後者葉綠素型態無法從原核生物或初級葉綠體得知,但是與紅藻基因相似性支持其關係存在。
属于色藻界的前三類,葉綠體有4層膜,隐藻殘留有類核體,他們很可能共同擁有含有色素的祖先。然而其他證據質疑这种说法,不等鞭毛類、定鞭藻和隐藻之間的關係是否比和其他類的關係更接近[10][11]。 典型甲藻類的葉綠體有3層膜,但是其葉綠體非常多樣,且顯示曾有數次內共生事件[12]。顶复门,一组關係十分接近的寄生生物,也有稱為顶复体 (Apicoplast)的色素體。顶复体無法行光合作用,但是顯示出和甲藻類葉綠體有共同起源[12]。 |
参考文献
- Butterfield, N. J. . Paleobiology. 2000, 26 (3): 386–404. ISSN 0094-8373. S2CID 36648568. doi:10.1666/0094-8373(2000)026<0386:BPNGNS>2.0.CO;2. (原始内容存档于7 March 2007).
- T.M. Gibson. . Geology. 2018, 46 (2): 135–138 [2024-03-13]. Bibcode:2018Geo....46..135G. doi:10.1130/G39829.1. (原始内容存档于2022-11-14).
- Paul, Vishal; Chandra Shekharaiah, P. S.; Kushwaha, Shivbachan; Sapre, Ajit; Dasgupta, Santanu; Sanyal, Debanjan. . Deb, Dipankar; Dixit, Ambesh; Chandra, Laltu (编). . Smart Innovation, Systems and Technologies 161. Singapore: Springer. 2020: 257–265 [2024-03-13]. ISBN 978-981-329-578-0. S2CID 202902934. doi:10.1007/978-981-32-9578-0_23. (原始内容存档于2023-04-06) (英语).
- Round, F.E. 1981. The Ecology of Algae. Cambridge University Press, London. ISBN 978-0-521-22583-0
- . [2008-08-14]. (原始内容存档于2008-03-21).
- Biology 8th ed. Losos, Jonathan B., Mason, Kenneth A., Singer, Susan R., McGraw-Hill. 2007.
- . [2008-10-14]. (原始内容存档于2021-04-03).
- Dixon, P S. . Edinburgh: Oliver & Boyd. 1973: 232. ISBN 978-0-05-002485-0.
- Bhattacharya, D.; Medlin, L., (PDF), Plant Physiology, 1998, 116 (1): 9–15 [2009-02-26], (原始内容存档 (PDF)于2009-02-07)
- Burki F, Shalchian-Tabrizi K, Minge M, Skjæveland Å, Nikolaev SI; et al. . PLoS ONE. 2007, 2 (8: e790): e790. doi:10.1371/journal.pone.0000790.
- Laura Wegener Parfrey, Erika Barbero, Elyse Lasser, Micah Dunthorn, Debashish Bhattacharya, David J Patterson, and Laura A Katz. . PLoS Genet. December 2006, 2 (12): e220 [2009-02-26]. PMID 17194223. doi:10.1371/journal.pgen.0020220. (原始内容存档于2019-09-12).
- Patrick J. Keeling. . American Journal of Botany. 2004, 91: 1481–1493 [2009-02-26]. doi:10.3732/ajb.91.10.1481. (原始内容存档于2008-02-27).
外部鏈接
- 陳衍昌藻類網址 (页面存档备份,存于) - 國立台灣海洋大學陳衍昌副教授藻類網址
- Algaebase - Algaebase
- 臺灣海藻資訊網 - 國立臺灣博物館
- 國立彰化師範大學生物系 - 王瑋龍藻類研究室 - 藻類查詢