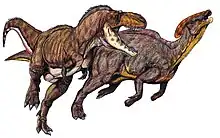
副櫛龍屬
副櫛龍屬、副龍櫛龍屬或似櫛龍屬(屬名:Parasaurolophus,意為“近冠蜥蜴”)[2],是鴨嘴龍科恐龍的一屬,是鸭嘴龙科里面发现化石最多、最具代表性的一个,生存於約7,650萬年到7,300萬年前的晚白堊紀[3],在美國和加拿大被大量發現化石。副櫛龍是種植食性恐龍,可以二足或四足方式行走,连接鼻头一直到后脑勺的头冠为其标志性特征。目前已有三個被承認種:模式種沃克氏副櫛龍(P. walkeri)、小號手副櫛龍(P. tubicen)以及短冠飾的短冠副櫛龍(P. cyrtocristatus)。
| 副櫛龍屬 | |
|---|---|
 | |
| 短冠副櫛龍(P. cyrtocristatus)骨骼化石,現存菲爾德自然史博物館 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | †鸟臀目 Ornithischia |
| 亚目: | †鸟脚亚目 Ornithopoda |
| 科: | †鴨嘴龍科 Hadrosauridae |
| 亚科: | †賴氏龍亞科 Lambeosaurinae |
| 族: | †副櫛龍族 Parasaurolophini |
| 属: | †副櫛龍屬 Parasaurolophus Parks, 1922 |
| 模式種 | |
| † 沃克氏副櫛龍 Parasaurolophus walkeri Parks, 1922 | |
| 物種 | |
 | |
| 異名[1] | |
| |
副櫛龍的首次敘述是在1922年,由威廉·帕克斯(William Parks)藉由在亞伯達省發現的一個頭顱骨與部分骨骸敘述。副櫛龍為常見的鴨嘴龍類,但目前良好标本为少數,化石發現於加拿大亞伯達省、美國的新墨西哥州與猶他州。副櫛龍因牠們的頭蓋骨上大型、修長的冠飾著名,冠飾往頭後方彎曲。副櫛龍的最親近物種應是最近在中國新發現的卡戎龍,兩者的顱骨類似,可能具有相似的冠飾。這種結構引起許多科學文獻的討論;現在對於該冠飾主要功能的意見包括:辨別性別與物種、共鳴器、以及調節體溫。
副櫛龍最先被認為與櫛龍(有冠飾的蜥蜴)是近親,但并非如此,櫛龍反而是和没有头冠的爱德蒙托龙相近。
敘述

副櫛龍只發現過部分身體骨骸。沃克氏副櫛龍的模式標本身長10-12公尺,頭顱骨與冠飾長1.6公尺。小號手副櫛龍模式標本的頭顱骨與冠飾超過2公尺,顯示牠們比沃克氏副櫛龍大[4]。副櫛龍重達7噸[5]。從目前唯一發現的前肢顯示,牠們的前肢比其他鴨嘴龍科恐龍的前肢短,並擁有短而寬的肩胛骨。沃克氏副櫛龍的模式標本股骨長達103公分,而且股骨粗壯[6]。副櫛龍上臂與骨盆都很粗壯[7]。
如同其他鴨嘴龍類,副櫛龍是二足恐龍,但可以轉換成四足行走。副櫛龍可能在尋找食物時採用四足方式,而在奔跑時採用二足方式[8]。副櫛龍脊椎上的神經棘高大,這特徵常見於賴氏龍亞科恐龍[6],這特徵增加背部高度,超過臀部的高度。已發現沃克氏副櫛龍的皮膚痕跡,顯示皮膚上有瘤狀鱗片[9]。
副櫛龍最著名的特徵是頭頂上的冠飾,由前上頜骨與鼻骨所構成,從頭部後方延伸出去。在沃克氏副櫛龍模式標本的脊椎上,一個可能是冠飾接觸到背部的地方,神經棘上有個凹口,但這可能是該個體的病理[6]。替副櫛龍命名的威廉·帕克斯(William Parks),假設從冠飾到脊椎凹口有個韌帶用來支撐頭部[9],但這似乎不太可能[5]。在許多重建模型裡,冠飾到頸部則是有塊皮膜。
副櫛龍的冠飾是中空的,內部有從鼻孔到冠飾尾端,再繞回頭後方,直到頭顱內部的管。沃克氏副櫛龍的管最簡單,而小號手副櫛龍的管最複雜,有些管是不通的,而其他管是交叉、分開的[10]。沃克氏副櫛龍、小號手副櫛龍的冠飾較長、微彎,而短冠副櫛龍的冠飾較短[11]。
分類

最初,副櫛龍被認為跟櫛龍有親緣關係,因為牠們的冠飾外型相似[9]。但是不久後,副櫛龍重新被歸類於賴氏龍亞科[12]。副櫛龍常被認為是賴氏龍的支系,不同於有頭盔狀冠飾的冠龍、亞冠龍、賴氏龍[8][13][14]。與副櫛龍最親近的屬是發現於中國東北黑龍江地區的卡戎龍,卡戎龍與副櫛龍的冠飾非常相似[15][14]。短冠副櫛龍的較圓、短冠飾,顯示牠可能是副櫛龍三個種最基礎、原始的一種[14],或者是小號手副櫛龍的亞成年個體或雌性個體[16]。
發現與命名
副櫛龍的模式標本(編號ROM708)包含一個頭顱骨與部分骨骸,缺少膝蓋以下的後肢與大部分的尾巴。該標本是在1920年,由一個多倫多大學的野外隊伍在加拿大亞伯達省紅鹿河畔的桑德河附近所發現[9]。這個標本的發現地點目前為恐龍公園層,年代為上白堊紀的坎帕階;而該化石被威廉·帕克斯命名為沃克氏副櫛龍(P. walkeri),以皇家安大略博物館的董事會主席Byron Edmund Walker爵士為名[9]。儘管在亞伯達省發現了副櫛龍的第一個標本,但牠們的化石在該省仍然少見[17]。除了模式標本以外,在恐龍公園組另發現了一個部分顱骨[18]、以及三個缺少顱骨的標本,可能屬於副櫛龍[19]。在蒙大拿州的地獄溪組,發現了疑似沃克氏副櫛龍的化石,該地層的年代為馬斯垂克階晚期[20][21]。這些未經處理過的蒙大拿州化石,是蘇利文、威廉森在1999年重新研究副櫛龍時發現的[10]。


在1921年,大約是亞伯達省的化石被發現、命名的同時,查爾斯·斯騰伯格(Charles Hazelius Sternberg)在新墨西哥州聖胡安縣的基特蘭德組發現了一個部份頭顱骨,該地層較恐龍公園組年輕。這個化石被送到瑞典烏普薩拉,卡爾·維曼(Carl Wiman)在1931年將牠們敘述成第二個種,小號手副櫛龍(P. tubicen)[22]。種名tubicen衍化自拉丁語中的吹鼓手[23]。小號手副櫛龍的第二個接近完整頭顱骨(編號NMMNH P-25100)是在1995年於新墨西哥州發現。在1999年,羅伯特·蘇利文(Robert Sullivan)與湯瑪斯·威廉森(Thomas Williamson)使用電腦斷層掃描來檢驗這個頭顱骨,並在一個專題論文上討論小號手副櫛龍的生理結構、分類,以及冠飾的功能[10]。湯瑪斯·威廉森稍晚提出了一個獨立的研究,提出不同的分類結論[16]。
在1961,約翰·奧斯特倫姆(John Ostrom)敘述了另一個保存良好標本,並命名為短冠副櫛龍(P. cyrtocristatus)。這個標本包含一個部份頭顱骨,上有圓、短冠飾,以及頭顱後的大部分骨骸,除了足部、頸部、以及部分尾巴以外;該標本目前位於菲爾德自然歷史博物館[11]。短冠副櫛龍的種名cyrtocristatus在拉丁語中意為「變短的冠飾」[23]。短冠副櫛龍被發現於Fruitland組頂層,以及其上的Kirtland組底層[10]。
在1979年,大衛·威顯穆沛(David B. Weishampel)與詹姆斯·詹森(James A. Jensen)在猶他州加菲爾德縣的凱帕羅維茨組發現了一個副櫛龍的部份頭顱骨,該地層年代為坎潘階[24]。這個頭顱骨與另一個猶他州所發現的頭顱骨,就冠飾的形態上屬於短冠副櫛龍[10]。
種
沃克氏副櫛龍(P. walkeri)是模式種,化石是單一個標本,發現於亞伯達省的恐龍公園層[8]。在恐龍公園組發現的其他標本,可能也屬於此種[3]。沃克氏副櫛龍與小號手副櫛龍的差別在於構造較簡單的冠飾內管[10];與短冠副櫛龍的差異則在於較長、較不圓的冠飾,以及肱骨比橈骨長[11]。沃克氏副櫛龍的生存年代約7650萬到7530萬年前[3]。
小號手副櫛龍(P. tubicen)的化石發現於新墨西哥州,化石來自於至少三個個體[8]。與沃克氏副櫛龍相比,小號手副櫛龍的冠飾較為複雜;與短冠副櫛龍相比時,小號手副櫛龍的冠飾較大,型狀也不一樣[10]。小號手副櫛龍的化石被發現於科特蘭層(Kirtland Formation)的De-na-zin段[25],地質年代約7340萬到7300萬年前,是生存年代最晚的物種[3]。
短冠副櫛龍(P. cyrtocristatus)的化石發現於新墨西哥州與猶他州,包含三個可能的標本。短冠副櫛龍是最小的副櫛龍,冠飾較短、較圓[10]。最近,湯瑪斯·威廉森根據短冠副櫛龍的體型小、冠飾形狀,提出一個假設,認為牠們是小號手副櫛龍的幼年體或雌性個體;而小號手副櫛龍也發現於新墨西哥同一地層,年代大約晚了一百萬年。威廉森注意到,短冠副櫛龍的體型大約是小號手副櫛龍的72%,這比例接近於賴氏龍亞科冠飾因為兩性異形而產生的不同大小(成年個體的70%)[16]。但是最近的賴氏龍亞科研究否定了這個論點[8][14]。短冠副櫛龍的化石被發現於凱佩羅維茲層(Kaiparowits Formation)與弗鲁特蘭層(Fruitland Formation)地層,地質年代約7550萬到7450萬年前[3]。
整體而言,目前有六個部份與完整的副櫛龍頭顱骨被發現:其中一個發現於亞伯達省(沃克氏副櫛龍)、兩個發現於猶他州(短冠副櫛龍)、以及三個發現於新墨西哥州(兩個是小號手副櫛龍,另一個是短冠副櫛龍)[10]。
古生態學
沃克氏副櫛龍的化石發現於恐龍公園層,該地層有許多保存良好且多樣性的史前動物群化石,包含許多著名的恐龍,例如:角龍科的尖角龍、戟龍、開角龍;鴨嘴龍類的原櫛龍、格里芬龍、冠龍、賴氏龍;暴龍科的蛇髮女怪龍;以及甲龍科的埃德蒙頓甲龍、包頭龍、倍甲龍[21]。副櫛龍在該動物群中很少見[17]。恐龍公園組被認為是河流與氾濫平原之間的低地,隨者時間經過,西部內陸海道越往西方海侵,而恐龍公園組變得更類似沼澤、更受到海洋環境的影響[26]。該地的氣候比今日的亞伯達省更為溫暖,無霜,但有更明顯的乾、濕季節變化。針葉樹明顯地是該地區的優勢、頂層植物,而底層植物則由蕨類、樹蕨、以及被子植物所構成[27]。
恐龍公園組的某些少見鴨嘴龍類,例如副櫛龍,可能是群體遷徙經過該地區時,少數個體死在該地區[28]。這些少見物種可能棲息於高地,在地勢較高地區築巢或進食[28]。出現於緯度較北地區的副櫛龍、小貴族龍化石,也可能代表白堊紀晚期的北美洲,曾發生北部、南部動物群的交流現象[29]。副櫛龍、小貴族龍、以及五角龍的大部分化石都發現於北美洲南部,牠們是南部的優勢草食性恐龍[29]。
而發現於新墨西哥州與猶他州的短冠副櫛龍,則是與以下恐龍共同生存者:角龍類的五角龍、猶他角龍、華麗角龍[30],虛骨龍類的似鳥龍,以及暴龍科的怪獵龍[31]。
體型最大、生存年代最晚的小號手副櫛龍,則是在新墨西州與以下恐龍共同生存者:大型的蜥腳類阿拉莫龍、鴨嘴龍類的小貴族龍、角龍類的五角龍、甲龍類的結節頭龍、蜥鳥盜龍、以及暴龍科的虐龍[21]。基德蘭組地層被認為是河流氾濫平原,應為西部內陸海道的海降痕跡。該地的優勢植物也是針葉樹,而角龍亞科比鴨嘴龍科更為常見[32]。
古生物學
進食
如同其他鴨嘴龍科恐龍,副櫛龍是種大型、草食性恐龍,可採二足或四足方式行走,複雜的頭顱骨容許類似咀嚼的磨碎運動。副櫛龍的牙齒是不斷地生長、取代,牠們有數百顆牙齒,同一時間只會使用少部份牙齒。副櫛龍使用牠的喙狀嘴來切割植物,並送入嘴部兩旁的頰部。牠們的進食範圍約為離地面4公尺以上的範圍[8]。羅伯特·巴克(Robert Bakker)提出,賴氏龍亞科的喙狀嘴比鴨嘴龍亞科的狹窄,顯示副櫛龍與牠的近親進食時,比寬廣嘴部、缺乏冠飾的鴨嘴龍亞科還更具選擇性[33]。
冠飾

關於副櫛龍冠飾的功能有許多假設,但許多是不足採信的[5][34]。現在認為冠飾有數種功能:辨別物種與性別的視覺展示物、溝通用的揚聲器、以及調節體溫。目前不確定在冠飾與內部鼻管的演化過程中,哪種功能是最重要的[35]。
不同種、年齡階段的差異
當提到賴氏龍亞科時,一般認為副櫛龍的冠飾隨者年齡而改變,並是成年個體的兩性異形特徵。詹姆斯·霍普森(James Hopson)是最早敘述賴氏龍亞科的冠飾並加以區分的研究人員之一,他提出有小型冠飾的短冠副櫛龍,是小號手副櫛龍的雌性個體[36]。湯瑪斯·威廉森則認為短冠副櫛龍是小號手副櫛龍的未成年個體[16]。兩個假設都不被廣泛接受。因為目前僅發現六個頭顱骨,若發現新的化石,將有助於這些種彼此間的關係。湯瑪斯·威廉森提出無論如何,未成年的副櫛龍可能有小、圓形的冠飾,類似短冠副櫛龍的冠飾,而冠飾在接近成熟年齡時可能加快成長速度[16]。根據近期的一個研究,一個原先被歸類於賴氏龍的幼年顱骨,目前被改歸類為副櫛龍,從此標本可知幼年副櫛龍具有較小的冠飾。該冠飾由額骨延伸出、支撐,與成年個體的冠飾形狀相似,但較小。這個顱骨標本也顯示副櫛龍的冠飾成長模式,與冠龍、亞冠龍、賴氏龍…支系的冠飾成長模式並不一樣;部分原因是這三個賴氏龍亞科的冠飾中央有冠脊,而副櫛龍則是長棒狀冠飾[18]。
被否定的功能假設


許多早期的假設認為鴨嘴龍科是兩棲動物,因此認為副櫛龍是依靠水而生的動物,這個普遍的想法持續到1960年代。因此阿爾弗雷德·羅默(Alfred Sherwood Romer)假設這些冠飾是當作潛水呼吸器使用[37],馬丁·威爾法斯(Martin Wilfarth)則這些管狀冠飾是當作活動式的呼吸與食物收集器[38],查爾斯·斯騰伯格(Charles Mortram Sternberg)則認為牠們可避免水進入肺中[39],而內德·科爾伯特(Edwin Harris Colbert)則認為它們被當作空氣儲藏室,可在水中待更多時間[40]。
其他的假設則較集中於生理方面。威廉·帕克斯認為冠飾與脊椎之間有韌帶或肌肉附著,可協助支撐頭部,以及協助頭部動作[9]。Othenio Abel則假設這些冠飾是作為物種內打鬥時的武器[41]。而安吉拉·米爾納(Angela Milner) 則認為它們是種葉狀變流器,類似鶴鴕的頭冠[34]。其他的假設則認為冠飾是種儲藏器官;例如Halszka Osmólska認為它們儲藏鹽腺[42],而約翰·奧斯特倫姆(John Ostrom)則認為賴氏龍亞科的冠飾內有嗅覺上皮的延伸區域,可協助牠們的嗅覺,而不具有防禦的功能[43]。其中一個最獨特的意見,創造論學者杜安·吉什(Duane Gish)所提出,他認為這些冠飾儲藏者化學腺體,當遇到敵人時,牠們可噴出化學液體,類似今日的放屁甲蟲[44]。
以上假設大部份遭到否定[5]。例如,冠飾後端並沒有洞口可供浮淺。沒有長鼻的肌肉痕跡,而且副櫛龍已經擁有喙狀嘴,是否還需要長鼻?如果作為氣鎖,這些冠飾並不能防止水進入。如果冠飾作為空氣儲藏室,因為副櫛龍的體型大,所提供的效率並不大。其他有大型頭部的鴨嘴龍科恐龍,並不需要大型中空冠飾來作為頸部韌帶附著點[43]。
而且沒有一個理論可以解釋其他衍生的問題,例如:冠飾的形狀、與其他賴氏龍亞科恐龍的不同形狀冠飾為何擁有類似的功能、缺乏冠飾或硬組織的鴨嘴龍科恐龍如何擁有類似的功能、以及為何某些鴨嘴龍科恐龍擁有硬組織冠飾。這些衍生問題質疑了某些生理方面假設,例如:鹽腺、嗅覺[34],並顯示這些冠飾並沒有主要的功能。此外,對於賴氏龍亞科的鼻腔研究顯示嗅覺神經與感覺器官並不位於冠飾的鼻管部分,所以這些冠飾應不具有嗅覺功能[35]。
社交功能
社交與生理功能現在為最多人支持的理論,尤其是視覺辨認物與揚聲溝通器。作為一個大型物體,副櫛龍的冠飾可作為明確的視覺辨認物,與同時代的其他動物做區隔。鴨嘴龍科的鞏膜環與大型眼眶,顯示牠們有精準的視力與日間習性,冠飾這種視覺辨認物對於這群動物將非常重要。如果副櫛龍的冠飾到頸後有皮膚皺摺,根據這個理論,這些皮膚皺摺將是華麗、引人注目的[36]。如同其他賴氏龍亞科的頭顱骨,副櫛龍頭顱骨的形狀與大小可用來辨認物種(例如冠龍與賴氏龍)與性別間差異[35]。
發聲功能
然而,冠飾的外形與內部的複雜管道並未相符合,顯示這些內部空間擁有其他的功能[35]。卡爾·維曼(Carl Wiman)在1931年提出這些管道可發出聽覺信號,如同克朗號[22],而詹姆斯·霍普森與大衛·威顯穆沛則在70年代與80年代分別重申這個理論[36][45][46]。霍普森發現有證據顯示鴨嘴龍科擁有良好的聽覺;在副櫛龍的近親冠龍身上發現了一個修長的鐙骨,有大型空間容納耳槌,顯示牠們有敏感的中耳;而鴨嘴龍科的聽壺如鱷魚般修長,顯示牠們擁有發展良好的內耳[36]。威顯穆沛則認為沃克氏副櫛龍能夠製造48到240赫茲的音頻,而短冠副櫛龍則可以製造75到375赫茲的音頻。他根據鴨嘴龍科與鱷魚的相似內耳,提出成年鴨嘴龍科恐龍對高音頻較為敏感,例如幼年鴨嘴龍類所發出的聲音。根據威顯穆沛的說法,這與成體及幼體之間的溝通吻合[45]。
電腦模擬顯示小號手副櫛龍的內部管道比沃克氏副櫛龍的還複雜,可使電腦重建出冠飾所發出的可能聲音[47]。主要的管道可發出約30赫茲的音頻,但複雜的鼻竇結構則可控制聲音的高峰與低峰[48]。
參考資料
- Martin 2014.
- (英文)Liddell, Henry George and Robert Scott. . United Kingdom: Oxford University Press. 1980. ISBN 978-0-19-910207-5.
- (英文)Evans, D.C., Bavington, R. and Campione, N.E. (2009). "An unusual hadrosaurid braincase from the Dinosaur Park Formation and the biostratigraphy of Parasaurolophus (Ornithischia: Lambeosaurinae) from southern Alberta. (页面存档备份,存于)" Canadian Journal of Earth Sciences, 46(11): 791–800. doi:10.1139/E09-050
- (英文)Lull, Richard Swann; and Wright, Nelda E. . Geological Society of America Special Paper 40. Geological Society of America. 1942: 229.
- (英文)Glut, Donald F. . . Jefferson, North Carolina: McFarland & Co. 1997: 678-684. ISBN 0-89950-917-7.
- (英文)Richard Swann Lull and Wright, Nelda E. 1942. Hadrosaurian Dinosaurs of North America, Geological Society of America Special Paper 40, 209-213.
- (英文)Brett-Surman, Michael K.; and Wagner, Jonathan R. . Carpenter, Kenneth (ed.) (编). . Bloomington and Indianapolis: Indiana University Press. 2006: 135–169. ISBN 0-253-34817-X.
- (英文)Horner, John R.; Weishampel, David B.; and Forster, Catherine A. . Weishampel, David B.; Osmólska, Halszka; and Dodson, Peter (eds.) (编). 2nd. Berkeley: University of California Press. 2004: 438-463. ISBN 0-520-24209-2.
- (英文)Parks, William A. . University of Toronto Studies, Geology Series. 1922, 13: 1–32.
- (英文)Sullivan, Robert M.; and Williamson, Thomas E. . New Mexico Museum of Natural History and Science Bulletin, 15. Albuqueque, New Mexico: New Mexico Museum of Natural History and Science. 1999: 1–52.
- (英文)Ostrom, John H. . Journal of Paleontology. 1961, 35 (3): 575–577.
- (英文)Gilmore, Charles W. . Canada Department of Mines Geological Survey Bulletin (Geological Series). 1924, 38 (43): 29–48.
- (英文)Weishampel, David B.; and Horner, Jack R. . Weishampel, David B.; Osmólska, Halszka; and Dodson, Peter (eds.) (编). 1st. Berkeley: University of California Press. 1990: 534–561. ISBN 0-520-06727-4.
- (英文)Evans, David C.; and Reisz, Robert R. . Journal of Vertebrate Paleontology. 2007, 27 (2): 373–393 [2007-07-28]. doi:10.1671/0272-4634(2007)27[373:AAROLM]2.0.CO;2.
- (英文)Godefroit, Pascal; Shuqin Zan; and Liyong Jin. . Compte Rendus de l'Academie des Sciences, Paris, Sciences de la Terre et des planètes. 2000, 330 (12): 875–882. doi:10.1016/S1251-8050(00)00214-7. </ref}}
- (英文)Williamson, Thomas E. . Lucas, S.G.; and Heckert, A.B. (eds.) (编). . New Mexico Museum of Natural History and Science Bulletin, 17. Albuqueque, New Mexico: New Mexico Museum of Natural History and Science. 2000: 191–213.
- (英文)Ryan, Michael J.; and Evans, David C. . Currie, Phillip J., and Koppelhus, Eva (eds.) (编). . Bloomington: Indiana University Press. 2005: 312-348. ISBN 0-253-34595-2.
- (英文)Evans, David C.; Reisz, Robert R.; and Dupuis, Kevin. . Journal of Vertebrate Paleontology. 2007, 27 (3): 642–650.
- (英文)Currie, Phillip J; and Russell, Dale A. 2005. "Geographic and stratigraphic distribution of dinosaur remains" in: Currie, Phillip J., and Koppelhus, Eva (eds.) Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed, p. 553.
- (英文)Weishampel, David B. (1990). "Dinosaur Distribution", in The Dinosauria (1st), pp. 63–139.
- (英文)Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loeuff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth, M.P.; and Noto, Christopher R. (2004). "Dinosaur Distribution", in The Dinosauria (2nd), pp. 517–606.
- (德文)Wiman, Carl. . Nova Acta Regia Societas Scientarum Upsaliensis, series 4. 1931, 7 (5): 1–11.
- (英文)Simpson, D.P. 5. London: Cassell Ltd. 1979: 883. ISBN 978-0-304-52257-6.
- (英文)Weishampel, David B.; and Jensen, James A. . Journal of Paleontology. 1979, 53 (6): 1422–1427.
- Sullivan, R., Jasinsky, S., Lucas, S. and Spielmann, J. (2009). "The first 'lambeosaurin' (Dinosauria, Hadrosauridae, Lambeosaurinae) from the Upper Cretaceous Ojo Alamo Formation (Naashoibito Member), San Juan Basin, New Mexico: Further implications for the age of the Alamo Wash local fauna." Journal of Vertebrate Paleontology, 29(3): 188A.
- (英文)Eberth, David A. 2005. "The geology", in Dinosaur Provincial Park, pp. 54-82.
- (英文)Braman, Dennis R., and Koppelhus, Eva B. 2005. "Campanian palynomorphs", in Dinosaur Provincial Park, pp. 101-130.
- Tanke, Darren H.; and Brett-Surman, Michael K. . Tanke, Darren H.; and Carpenter, Kenneth (编). . Bloomington and Indianapolis, Indiana: Indiana University Press. 2001: 206–218. ISBN 0-253-33907-3.
- Lehman, T. M., 2001, Late Cretaceous dinosaur provinciality: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, pp. 310–328.
- "New Horned Dinosaurs of Utah Provide Evidence for Intracontinental Dinosaur Endemism" in Sampson et al. (2010)
- (英文)Thomas D. Carr, Thomas E. Williamson, Brooks B. Britt and Ken Stadtman. . Naturwissenschaften. 2011, 98 (3): 241–246. PMID 21253683. doi:10.1007/s00114-011-0762-7.
- (英文)Russell, Dale A. . Minocqua, Wisconsin: NorthWord Press. 1989: pp. 160–164. ISBN 1-55971-038-1.
- (英文)Bakker, Robert T. . New York: William Morrow. 1986: 194. ISBN 0-8217-2859-8.
- (英文)Norman, David B. . . New York: Crescent Books. 1985: pp. 122–127. ISBN 0-517-468905.
- (英文)Evans, David C. . Paleobiology. 2006, 32 (1): 109–125 [2007-07-28]. doi:10.1666/04027.1.
- (英文)Hopson, James A. . Paleobiology. 1975, 1 (1): 21–43 [2007-07-28].
- (英文)Romer, Alfred Sherwood. . University of Chicago Press. 1933: p. 491. OCLC 1186563.
- (英文)Wilfarth, Martin. . Orion (Munich). 1947, 2: pp. 525–532 (德语).
- (英文)Sternberg, Charles M. . Canada Department of Mines Bulletin (Geological Series). 1935, 77 (52): 1–37.
- (英文)Colbert, Edwin H. . New York: American Museum of Natural History, Man and Nature Publications, 14. 1945: 156. OCLC 691246.
- (英文)Abel, Othenio. . Jarbuch Naturwissenschaften. 1924, 12 (36): 709–716 (德语).
- (英文)Osmólska, Halszka. . Acta Palaeontologica Polonica. 1979, 24: 205–215.
- (英文)Ostrom, John H. . Postilla. 1962, 62: 1–29.
- (英文)Gish, Duane T. . Green Forest: Master Books. 1992: p. 82. ISBN 0-89051-165-9.
- (英文)Weishampel, David B. . Paleobiology. 1981, 7 (2): 252–261 [2007-07-28].
- (英文)Weishampel, David B. . Journal of Paleontology. 1981, 55 (5): 1046–1057 [2007-07-28].
- (英文)Sandia National Laboratories. . 1997-12-05 [2007-06-30]. (原始内容存档于2014-08-17).
- (英文)Diegert, Carl F.; and Williamson, Thomas E. . Journal of Vertebrate Paleontology. 1998, 18 (3, Suppl.): 38A.
- (英文)Sullivan, Robert M.; and Williamson, Thomas E. . Journal of Vertebrate Paleontology. 1996, 16 (3, Suppl.): 68A.
- (英文)Wheeler, P.E. . Nature. 1978, 275: 441–443. doi:10.1038/275441a0.
- (英文)Maryańska, Teresa; and Osmólska, Halszka. . Lethaia. 1979, 12: 265–273.
外部連結
- Scientists Use Digital Paleontology to Produce Voice of Parasaurolophus Dinosaur; from Sandia National Laboratories.
- Restorations of P. walkeri and P. cyrtocristatus, to the same scale, by Scott Hartman; at Skeletal Drawing.com.
{kind=link}