華麗角龍屬
华丽角龙(属名:Kosmoceratops,发音:/ˌkɒzməˈsɛrətɒps/[1])是角龙科恐龙的一个属,生存于大约7600至7590万年前晚白垩世的北美。标本于2006及2007年在犹他州大升梯国家纪念区的凯帕罗维兹组发现,包括成年恐龙的颅骨和颅后骨骼及部分亚成体骨骼,其中前者于2010年成为新属新种里氏华丽角龙(Kosmoceratops richardsoni)的正模标本。属名意为“华丽的有角面孔”,种名纪念标本发现者斯科特·理查德森(Scott Richardson)。作为21世纪初众多角龙类新发现之一,华丽角龙因其复杂的颅骨装饰而被认为意义重大。
| 华丽角龙属 化石时期:晚白垩世, | |
|---|---|
 | |
| 含重建的前齿骨、右角及左脸的颅骨模型 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | †鸟臀目 Ornithischia |
| 科: | †角龙科 Ceratopsidae |
| 亚科: | †開角龍亞科 Chasmosaurinae |
| 属: | †华丽角龙属 Kosmoceratops Sampson et al., 2010 |
| 模式種 | |
| †里氏华丽角龙 Kosmoceratops richardsoni Sampson et al., 2010 | |
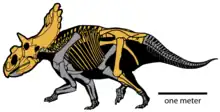
估计华丽角龙长4.5米(15英尺)、重1.2 t(1.3 short ton)。作为角龙科,华丽角龙是种身形笨重的四足动物,拥有尖锐的三角形喙嘴及末端扁平的刃状鼻角。在其大多数近亲中,眼睛上方的角皆指向前方或后方,而华丽角龙的角则向上指并向两侧倾斜,然后向下弯曲,最后以尖锐末端结束。颈盾长度较短,具有小型顶骨窗(穿透颈盾的开口),后缘有10个钩状突起,其中8个向前弯曲,2个侧向弯曲。颈盾上有15个发育良好的角和角状结构,使其拥有已知恐龙物种中最华丽的颅骨。华丽角龙隶属开角龙亚科,起初被认为与迷乱角龙(颈盾背面也有前弯的突起)关系密切,但一直存在争议,因为部分研究者发现后者与开角龙关系更近。鉴于颈盾类似,本属也被认为与钉盾龙关系较近。
骨组织学研究表明华丽角龙生长迅速,有强大的新陈代谢,与现代鸟类及哺乳类相似。角龙科牙齿适合处理纤维植物,凯帕罗维兹组含木屑的粪化石可能是由角龙科制造。角龙类颈盾装饰和角的功能一直存在争议,包括展示、战斗及种内识别。凯帕罗维兹组的地质年龄可追溯到坎帕阶晚期,沉积于拉腊米迪亚古陆――北美中部被西部内陆海道分开时形成的陆岛――上。其环境以湿地为主,繁育着多种动物群,包括开角龙亚科的犹他角龙等恐龙。考虑到华丽角龙与其它大约同时期出现的开角龙亚科之间的关系,有人指出,拉腊米迪亚被划分为许多具有孤立特有种区的“恐龙省”(表明南方的华丽角龙与北方地理隔离的迷乱角龙关系最近),但该假说一直存在争议。
发现

自2000年以来,犹他州自然历史博物馆(UMNH)和美国内政部土地管理局一直在犹他州南部大升梯国家纪念区的凯帕罗维兹组进行古生物考察。该国家纪念区于1996年建立,部分原因是为了保护和研究化石,现已出土大量该区域独有的恐龙。该项目被称为“凯帕罗维兹盆地项目”,其它机构的实地工作者亦参与其中。[2][3]项目取得的发现包括三种新的角龙类分类单元,其中之一出土自野外工作者斯科特·理查德森于2006及2007年野外季期间发现的两个地区(编号为UMNH VP 890和951),最初被称为“凯帕罗维兹新分类单元A”,后来于2010年一本有关角龙类的学术研讨会书中鉴定为开角龙亚科角龙科恐龙。[3][4]挖出的化石由直升机空运至UMNH(化石发现地),志愿者在该地点用风镐和针制备标本然后重新组装。[2][5]
2010年,古生物学家斯科特·桑普森(Scott Donald Sampson)、马克·洛温(Mark A. Loewen)、安德鲁·法克(Andrew Allen Farke)、艾里克·罗伯特茨(Eric M. Roberts)、凯瑟琳·弗斯特(Catherine Ann Forster)、约书亚·史密斯(Joshua A. Smith)和艾伦·提图斯(Alan L. Titus)正式命名新属新种里氏华丽角龙(Kosmoceratops richardsoni),并将UMNH VP 17000(发现于VP 890地区)选为正模标本。属名取自古希腊语κόσμος/kosmos(华丽的)、κέρας/keras(角)和ὤψ/oops(脸),其中ceratops为命名角龙类的常用后缀;种名致敬在大升梯纪念区发现正模标本及许多其它化石的理查德森。[2]全名可译作“理查德森的华丽有角面孔”。[1]正模标本包括一个近乎完整的成体颅骨,缺少前齿骨(下颌最前端的骨骼)及面部和颈盾左侧的一小部分(部分颧骨、鳞状骨和顶骨),鼻部也因死后变形而向右弯曲。已发现大部分与颅骨相关的中轴骨(如椎骨和肋骨)及部分骨盆和四肢骨骼。据认为有45%的颅后骨骼保存下来,截至2010年,其中大部分仍在制备中。[2][4][6]归入该属的标本还包括UMNH VP 16878[3]――一具约为成体一半大小的无关节亚成体(介于幼年和成年之间)颅骨,缺少前颌骨、鼻骨和前坐骨;以及UMNH VP 21339――一具脱节的亚成体或成体骨骼。据报道总共发现四件标本。[2][7][3]
描述者还在同一论文中命名开角龙亚科新属犹他角龙(也来自凯帕罗维兹组)和迷乱角龙(来自恐龙公园组,唯一种尔文迷乱角龙曾被归入开角龙),[2]两者与其所在分类群的典型成员相比非常独特,也是21世纪初众多角龙类新发现之一,当时有大量新分类单元被命名(2013年的一项研究表明,所有角龙科有效属中有一半是自2003起被命名,这十年被称为“角龙科复兴”)。[1][3][8]桑普森及其同事的文章主要关注三者如何为确认其生存时间和地点的古生物地理学提供新证据。[2]2010年宣布该研究的新闻稿中,桑普森将华丽角龙描述为“已知最神奇的动物之一,巨大的颅骨上装饰着各种骨质突起”,并指出大升梯纪念区是“美国最后一个基本未被开发的大型恐龙骨场之一”。[1]
2017年,美国政府宣布计划缩减大升梯(缩减略超过规模的一半)和熊耳国家纪念区以便在这片土地上进行煤炭开采和其他能源开发,这是美国历史上最大规模的国家纪念区缩减。[9][10]桑普森曾负责该纪念区的大部分早期研究,表示担心该举动会威胁进一步的发现。媒体强调该地区化石发现的重要性――包括超过25个新分类单元,有些媒体则强调华丽角龙是最重要的发现之一。[10][11][12][13]美国政府随后被科学家、环保主义者和美国原住民起诉,诉讼正在进行中。[9][12][14]
1928年,化石猎人查尔斯·斯腾伯格(Charles M. Sternberg)在加拿大阿尔伯塔省的恐龙公园组发现部分颅骨(编号CMN 8801),该颅骨于1940年归入罗氏开角龙,但在1995年因颈盾缺失导致无法鉴别物种而仅归入开角龙。2014年(还有2015年一篇同行评审失败的文章中),古生物学家尼古拉斯·朗里奇(Nicholas R. Longrich)认为该颅骨鼻部特征与华丽角龙相似,但鼻孔和鼻角形状不同,因此提出这是除里氏种外的另一个华丽角龙物种,并将其归入华丽角龙未定种。他指出现在命名新种还为时过早,因为通常需要颈盾来鉴别特定角龙科物种,况且目前只描述了一个颅骨,因此很难确定该物种(里氏种)的特征及变异范围。[6][15]2016年,古生物学家詹姆斯·坎贝尔(James A. Campbell)及其同事反对将标本CMN 8801归入华丽角龙,因为他们发现其所依据的特征要么受埋藏学(腐烂和石化过程中发生的变化)影响,要么属于开角龙标本的变异范围(尽管他们没有将其归入任何物种)。[16]古生物学家丹佛·福勒(Denver W. Fowler)和伊丽莎白·弗里德曼·福勒(Elizabeth A. Freedman Fowler)2020年称,当更好地了解开角龙亚科颅骨前部的结构时,可能对CMN 8801作出更可靠的分类。[17]
描述
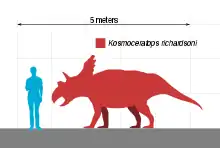
估计华丽角龙长4.5米(15英尺)、重1.2 t(1.3 short ton)。作为一种角龙科,它应该是种骨骼结构笨重、骨盆较大、尾部较短的四足动物。主齿列很长,形成由无齿喙嘴后边数百颗牙齿组成的复杂切割齿系。[18][2]上前颌喙呈三角形,最前面的喙骨外形尖锐,尖端向前并向下突出。鼻角角核(Horncore,鼻角生长处的骨质突起)呈刃状,底部从前到后伸长变窄,上端扁平,在开角龙亚科中很不寻常。鼻角核整体呈四边形,位于鼻部背面相对较远的位置。鼻孔(骨质鼻部开口)较高,前后宽度较窄,显然呈椭圆形而非近圆形,位置明显靠后,与其它开角龙亚科不同。前颌骨的鼻孔支(Narial strut,从鼻腔底部延伸至顶部)也向后倾斜(该特征也见于准角龙和无鼻角龙),从前颌骨向后向上突出的鼻骨突具有三角形尖锐末端。[2][6]

眼眶周围区域非常独特。大多数开角龙亚科的眶上角(眼睛上方的角)要么前倾,要么后倾,而华丽角龙和犹他角龙的角则指向上方并向两侧弯曲,类似现代野牛。与后者短而钝的角相比,前者更为细长,先向上指然后向下弯曲,最后形成尖锐末端。颅顶在眼眶正前方及中间有一明显突起,该突起亦存在于少数其它角龙科中,如恶魔角龙。华丽角龙拥有较大的上颧骨化物(Epijugal ossification,或称“颧角”),对开角龙以外的开角龙亚科来说非常典型。侧颞孔(颅骨两侧的孔)又高又窄。[2][6]
华丽角龙的顶鳞骨(Parietosquamosal)颈盾(由顶骨和鳞状骨形成)非常衍生(或“先进”),仅与迷乱角龙存在几个共有特征,包括颈盾很短,拥有位置非常靠后的小型顶骨窗(Parietal fenestrae,穿过褶边的开口),后缘存在十个钩状表层骨化物(Epiossification,形成角并排列在角龙科颈盾边缘的附属骨化物)。华丽角龙颈盾比迷乱角龙为极端:宽度约为长度的两倍(在块个骨骼表面测量),顶骨窗更小,位置更靠后,后缘有更细长而明显的表层骨化物。尽管开角龙亚科传统上被视为“长颈盾”角龙科,但华丽角龙颈盾长度与宽度之比在该亚科中最小,顶骨窗与颈盾总面积之比也是所有角龙科最小的。鳞状骨在侧视图中强烈弯曲,在俯视图中侧缘平行。顶骨没有突出到鳞状骨前方,V形后缘上的一排凹痕横贯整块骨骼。[2][6]
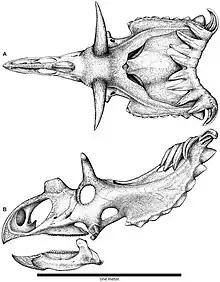
华丽角龙的鳞状骨上的表层骨化物沿颈盾背面逐渐增大。颈盾后缘每侧有五个发育良好的表层骨化扬:顶骨上有三个前弯的上顶骨(Epiparietal/ep1–3),顶骨和鳞状骨的边界上有一前弯的上顶鳞骨(Epiparietosquamosal/esp),鳞状骨上有一指向侧面及下方的上鳞骨(Episquamosal/es1)。前弯的上顶骨拥有底部融合的突出沟槽。华丽角龙有15个发育良好的角和角状结构,拥有已知恐龙中最“华丽”的颅骨,包括一个鼻角核、两个眶后角核、两个上颧骨及颈盾背面十个发育良好的表层骨化物。[2][6]亚成体标本UMNH VP 16878具有与成年正模标本相同的表层骨化物数量及模式,或许可以区分华丽角龙和犹他角龙的亚成体生长阶段。[3]
分类学
主要根据颅顶的复杂装饰来鉴别角龙科的两个演化支(所有拥有共同祖先的分类单元组成的分类群)――尖角龙亚科与开角龙亚科。鉴于前颌骨有鼻孔支和三角形突起及细长鳞片状骨的存在,桑普森和同事于2010年将华丽角龙归入后者。系统发育分析发现华丽角龙是迷乱角龙的姐妹群(在分支图中,两者形成的分支比无鼻角龙更原始、比科阿韦拉角龙更先进),两者处在由晚白垩世坎帕阶末期至马斯特里赫特阶衍生(包括三角龙,但与更基础(或“原始”)的开角龙关系较远的开角龙亚科组成的演化支中。[2]古生物学家乔丹·马龙(Jordan C. Mallon)和同事2011年的研究支持该结论,古生物学家斯蒂芬·威克(Steven L. Wick)和托马斯·勒曼(Thomas M. Lehman)2013年的研究也是如此。[19][20]
2011及2014年,朗里奇发现迷乱角龙是开角龙的一个种,符合最初分类(当时命名为文尔开角龙),而华丽角龙和其它分类单元之间没有过近的关系。[21][6]在2015年同行评审失败的文章中,朗里奇认为两者颈盾装饰的相似性是由趋同进化所致,指出尔文开角龙(或迷乱角龙,他当时承认其可能为独立属)颈盾背面的十个表层骨化物仅位于顶骨上,但华丽角龙的表层骨化物却位于顶骨和鳞骨上(由桑普森及其同事首次发现)。[15]坎贝尔在2014年的一篇硕士论文中指出,桑普森及其同事最初将成年华丽角龙颈盾后缘的全部十个表层骨化物皆鉴别为上顶骨,后来检查顶骨和鳞状骨之间骨缝更明显(随年龄增长,骨缝融合且变得模糊)的亚成体标本时才意识到其中只六个上顶骨。因此,桑普森和同事们亦就迷乱角龙表层骨化物模式给出同样的新解释,但坎贝尔认为该属有四至五个上顶骨,并发现它是开角龙而非华丽角龙的姐妹群。[22]2016年,坎贝尔和同事还发现,与华丽角龙相比迷乱角龙和开角龙的关系更近,[16]并2019年发现迷乱角龙是开角龙的一个种。[23]
古生物学家丹佛·福勒(Denver W. Fowler)和同事于2011年的一次会议摘要中提出,角龙科有很多属仅为其它属的不同个体发育形态(或生长阶段),而华丽角龙实际上是尔文开角龙的未成熟阶段(明显的颅骨装饰随年龄增长而减少)。[24]2015年,古生物学家卡雷布·布朗(Caleb M. Brown)和唐纳德·亨德森(Donald M. Henderson)测试不同开角龙亚科(各分类单元之间互相对应)表层骨化物之间同源的新方案,并发现包含华丽角龙和犹他角龙的演化支是所有其它开角龙亚科的姐妹群,与早期研究相反。[25]古生物学家格雷戈里·保罗(Gregory S. Paul)在2016年的一本畅销书中指出,华丽角龙和迷乱角龙与开角龙的差异不足令其独立建属,而应将前两者皆归入开角龙(或将迷乱角龙并入华丽角龙),并将开角龙亚科其它几个属列为开角龙的异名。[18]角龙科大部分属一般被视为单型属(仅含一个物种)。[26]


2016年,马龙和同事发现华丽角龙和迷乱角龙形成演化支,新属钉盾龙为其姐妹群。尽管承认一些研究者将迷乱角龙与开角龙归入同一演化支,但他们发现颈盾后面的前弯表层骨化物非常独特,显示钉盾龙可能为解释该特征的演化提供线索。他们认为,表层骨化物的弯曲是在向华丽角龙进化的谱系上逐步发生,首先是中间弯曲,然后是向两侧弯曲。由于在钉盾龙化石上处于更“原始”的停滞状态,因此该过程可能发生在颈盾缩短及顶骨窗缩小之前。尽管钉盾龙在该方面疑似处于过渡状态,但与华丽角龙生活在相同时期,因此不可能是其祖先。以下分支图遵循马龙和同事2016年的分析:[27]
| 角龙科 Ceratopsidae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
2020年,福勒和弗里德曼·福勒描述开角龙亚科的两个新属,并指出该亚科在包含开角龙及其近亲和五角龙及其近亲的演化支间存在高度进化分裂。与之前的研究相反,华丽角龙和迷乱角龙皆被发现与开角龙关系更近,代表该谱系中最原始兼最年轻的成员。根据该解释,华丽角龙是从迷乱角龙进化而来,而迷乱角龙是从开角龙进化而来。由于发现迷乱角龙可能是华丽角龙的姐妹群,他们建议将其作为与开角龙不同的属加以保留,因为在更详细地了解开角龙之前,其位置可能会长期不稳定。该研究未对钉盾龙和其它近期命名的开角龙亚科物种进行评估。[28]
古生物学

2013年的一篇硕士论文(总结于2019年发表的论文中[29])中,古生物学家卡罗琳·盖尔·莱维特(Carolyn Gale Levitt)对华丽角龙(成年正模标本股骨及归入该属亚成年或成年标本UMNH VP 21339)和犹他角龙的长骨进行组织学研究,以检视骨显微结构的生长及成熟标志(在此之前,只对开角龙亚科做过这类研究)。骨组织内有大量骨细胞和密集的血管网,包括辐向血管沟(向骨内部延伸的血管沟),表明其持续快速生长。这些特征还表明角龙类新陈代谢较高,像现代鸟类和哺乳类一样属于恒温吸热动物(或“温血动物”)。莱维特采样的华丽角龙和犹他角龙骨骼并未显示生长停滞线(年度生长线)的证据,与来自更远北方且具有生长线的厚鼻龙、尖角龙和野牛龙相比,可能表明气候会影响骨骼生长,而华丽角龙和犹他角龙因南方气候更平均而全年皆可持续生长。缺乏生长线也表明无法估计华丽角龙和犹他角龙标本的个体年龄,但莱维特确定被检查的华丽角龙标本是亚成体或成体,并推翻福勒及其同事2011年的结论,即两者皆为未成熟的迷乱角龙。她还确认最大的华丽角龙和犹他角龙标本都是成年个体,因此排除其中一只是另一只的幼龙的可能。[7]
2017年的一篇硕士论文中,古生物学家尼可·玛丽·瑞格韦尔(Nicole Marie Ridgwell)描述凯帕罗维兹组发现的两种粪化石,从大小来看,可能是由地层中已知三种植食恐龙类群之一的成员所制造:角龙类(包括华丽角龙)、鸭嘴龙类或甲龙类(三者中最为罕见)。粪化石含被子植物木材碎片(表示恐龙食用了木本植物),虽然以前少有证据表明恐龙食用被子植物,但这些粪化石显示恐龙已适应于以被子植物(首次常见于早白垩世,在晚白垩世呈现多样化)为食。粪化石中还出现过软体动物外壳、节肢动物角质层和蜥蜴骨骼的痕迹,是在其它含松柏门木屑的植食动物粪化石中发现,这些异物可能是与植物材料一起摄入体内。瑞格韦尔指出,角龙类和鸭嘴龙类的牙齿解剖结构(齿系由替换齿组成)适合处理大量纤维植物。粪化石代表的不同饮食可能表明了凯帕罗维兹组生态系统植食动物间的生态位分化或饮食存在季节性变化。[30]
颅骨装饰的功能

尽管主流假说涉及提高繁殖成功率,但角龙类的角和颈盾的可能功能仍存在争议,包括抵御捕食者、种内识别和调节体温。桑普森在2010年的一篇新闻稿中介绍华丽角龙、犹他角龙和迷乱角龙,表示这些“奇异特征”中的大多数皆不足以对付掠食者,但会被用来恐吓或攻击同性竞争对手并吸引异性。[1]2011年,古生物学家凯文·帕迪安(Kevin Padian)和约翰·霍纳(John R. Horner)提出,恐龙的“奇异结构”(包括角、颈盾、圆顶和冠)主要用于种内识别(区分共域种或同期共存的近缘种),并驳斥其它没有证据支持的假说。他们指出,许多大型角龙类的颈盾上都有开口,因此在防御时几乎没有用处,而角的大小及倾斜方向的多样性在战斗中也没有明显作用。他们还指出,几乎没有证据表明角龙类存在两性异形。[31]同年,古生物学家罗布·克内尔(Rob J. Knell)和桑普森在反驳帕迪安和霍纳时指出,虽然种内识别可能是“奇异结构”的次要功能,但用于性选择(用于展示或争夺配偶)的可能性更大,因为长出这类结构的成本很高,而且种内差异也很大。他们还指出,缺乏两性异形不排除在交配竞争中使用角:雄牛的角就是用于此目的,而雌牛的角主要用作防御,其次是性选择。[32]
2013年,古生物学家大卫·宏恩(David E. Hone)和德恩·耐许(Darren Naish)批评了“种内识别假说”,认为没有现存动物主要将这类结构用于种内识别,而帕迪安和霍纳忽视了相互性选择的可能性(两性都有装饰)。他们指出,如果这些结构的主要功能是种内识别,那么单个结构形状就会存在差异,因为额外成本会使额外结构变得多余。例如,角龙类有复杂的鼻角、眉角、颧骨突、颈盾中线和边缘特征及身体大小和比例的差异,而缺少或多出一支角就足以区分共域种。[33]2018年,古生物学家安德鲁·克纳普(Andrew Knapp)和同事研究角龙类的不同装饰性特征是否像“种内识别假说”所预测的那样与两个或多个物种之间同域共存有关。他们发现这些装饰的结构总体上差异很大,但在共域种与非共域种之间并没有显著差异,从而得出结论称该假说在角龙类中没有统计支持。[34]
2015年,生物学家帕斯夸尔·拉亚(Pasquale Raia)及其同事以菊石、鹿和角龙类为例,研究动物装饰(如冠、角和长牙)的复杂性及尺寸随时间增加的演化。较晚的物种中,角龙类颈盾复杂性似乎有所增加,而华丽角龙颈盾边缘的分形维数值最高(其次是戟龙、恶魔角龙和尖角龙)。作者发现装饰复杂度随体型增大而增大,表明其进化是科普法则(该法则假设种群谱系的体型随进化时间推移而增大)的副产物。他们认为,尽管大多数装饰物的尺寸可能受性选择控制(以及它们是否用于选择配偶、竞争或两者兼有),但并不影响其复杂性及外观,并得出结论:性选择可能影响装饰的尺寸而非形状。[35]
古生态学

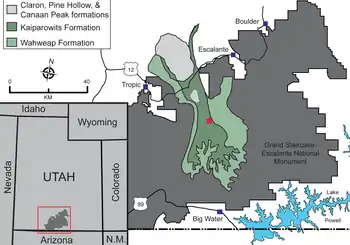
华丽角龙生存于犹他州的凯帕罗维兹组,地质年龄可追溯到晚白垩世坎帕阶后期,化石发现于地层下层上段至中层上段7640至7550万年前的沉积物中。该地层沉积在距西部内陆海道(分隔北美中部的内海)不出100(62英里)的拉腊米迪亚大陆(由当今北美西部构成的陆岛,其东面的大陆称作“阿巴拉契亚”)东岸一盆地(即西部内陆盆地)的南部。[2][36][37]该盆地平坦宽阔,呈新月形,除东边的西部内陆海道外皆被山脉包围。[38]该地层代表温热潮湿的冲积层至沿海平原环境,主要由河岸线稳定的大型河道和常年的湿地沼泽、池塘及湖泊组成。其河流一般向西流经平原汇入西部内陆海道,美国墨西哥湾沿岸地区被视为与之非常相似的现代地区(例如现在的路易斯安那沼泽地)。该地层保存了丰富多样的化石,包括陆生及水生动物、植物和孢粉(有机微体化石)。[39][5]

凯帕罗维兹组的其它鸟臀目恐龙包括角龙类如开角龙亚科的犹他角龙及尖角龙亚科的大鼻角龙(可能还有第二种未命名尖角龙亚科)、分类不明的厚头龙类、甲龙科的钉头龙、分类不明的结节龙科、鸭嘴龙类的钩鼻龙和副栉龙及分类不明的基干新鸟臀类。兽脚类包括暴龙科的怪猎龙、偷蛋龙类的哈格里芬龙、未命名的似鸟龙科、伤齿龙科的塔罗斯龙、分类不明的驰龙科及鸟类的鸟龙鸟。其它脊椎动物包括鳄鱼如恐鳄和短鳄、海龟如阿杜库斯龟属和皇室龟、翼龙、蜥蜴、蛇、两栖类、哺乳类及鱼类。[38][40][41]地层中最常见的两类大型脊椎动物是鸭嘴龙类和角龙类(后者约占该所有脊椎动物化石的14%),可能表明两者在凯帕罗维兹动物群中非常丰富,也可能反映出埋藏偏差(一种采样偏差),因为两者拥有最坚固的骨骼材料。[4]地层还发现了恐龙、鳄鱼和海龟的蛋。[42]沼泽和湿地主要生长高达30(98英尺)的柏树、蕨类和包括巨型浮萍、水浮莲及其他漂浮被子植物在内的水生植物。稍干燥的地区主要生长高达10至20米(33至66英尺)的双子叶植物、少量棕榈树和包括蕨类在内的下层植被。远离潮湿区域的干燥区主要生长高达30(98英尺)的针叶林和包括苏铁、小型双子叶树木或灌木丛可能还有蕨类在内的下层植被。[38]
埋藏学

2010年,古生物学家迈克尔·盖蒂(Michael A. Getty)和同事研究正模标本和亚成体标本UMNH VP 16878的埋藏学及保存标本的沉积环境。正模标本上的关节几乎全部连接,而且是在泥质砂岩河道岩相(沉积环境的岩石记录)中发现,与尸体被冲入河道再迅速掩埋的情况一致。尸体沉积之前,四肢及部分尾巴可能已经腐烂或被食腐动物破坏。颅骨左侧缺失部分在被发现以前因风化而缺失。由于当时大部分骨骼仍在制备中,研究者无法检查上面是否有被掠食者或被食腐动物咬过的很迹。亚成体标本UMNH VP 16878的骨骼被发现分散于3平方(32平方英尺)的区域内,骨骼脱节严重及断裂部分表明,该标本在埋进泥质泥岩岩相前就已经变成骷髅并且腐烂。[4][2]莱维特报告称,归入该属的亚成年或成年标本UMNH VP 21339的每一块骨骼似乎皆已在沉积前断裂,其基质由粉砂岩和泥岩及少量砂岩堆叠而成,表明当时存在池塘环境。[7]
古生物地理学
.jpg.webp)
虽然拉腊米迪亚古陆的面积仅有现代北美的20%,但其见证了恐龙的主要演化辐射,包括常见鸭嘴龙类及角龙类。据推测,在晚白垩世坎帕阶及马斯特里赫特阶的拉腊米迪亚岛上存在数个按纬度排列的“恐龙省”或生物群落,边界位于现在的犹他州北部至科罗拉多州一带;大陆南北皆存在相同的主要演化支,但所含属种不同。该假说受到了挑战:一种观点认为,该时期的南北恐龙群落并未同时存在,而是反映出一种随时间推移的分类学分布,以给人存在地理孤立“省份”的假象;不同群落可能是地层间采样偏差的产物。由于拉腊米迪亚南部缺乏可精确定年的化石,因此该猜想很难验证,但凯帕罗维兹组的发现增加了对该地区晚白垩世脊椎动物化石的认识。角龙科演化辐射似乎在时间和地理分布上皆非常有限(物种更替率极高,每个物种的存在时间都不到100万年),多数分类单元皆发现于西部内陆盆地的晚白垩世沉积物中,因此可能是在拉腊米迪亚起源并走向多样化。[2][43]
华丽角龙和犹他角龙的发现大大增加了西部内陆盆地已知开角龙亚科的数量。桑普森和同事认为这是对2010年描述中“恐龙省”概念最有力的支持,并指出,与马斯特里赫特阶相比,之前的坎帕阶具有更好的标本、多样性和范围更广的恐龙群及更精确的地理与地层数据。地层延续时限(Stratigraphic range)表明华丽角龙和犹他角龙生存于相同时期,且可能生活在同一生态系统中,这在角龙类中非常罕见。根据桑普森及其同事的研究,地质年代学定年显示,凯帕罗维兹组时限短暂的地质层段与阿尔伯塔的恐龙公园组及北部(上朱迪思河组和双麦迪逊组)和东南(果园组和阿古哈组)的其它地层年代相同、时限短暂且时期互相重叠,这些地层的纬度跨距允许各自之间进行重要比较。桑普森和同事指出,华丽角龙和犹他角龙彼此之间――或与同时期恐龙公园组之开角龙和魅惑角龙及阿尔伯塔之迷乱角龙和同时出现的新墨西哥之五角龙――关系并不密切,并以此为证据反驳该时期南北恐龙群生活在不同时期的假说。[2]
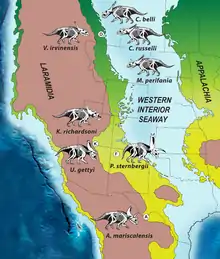
桑普森和同事称其研究首次记录到恐龙的陆内特有种分布(不同开角龙亚科分类单元在拉腊米迪亚的南北部同时存在),并发现开角龙亚科进化历程中的一系列事件与系统发育、地层和生物地理学证据一致。根据他们的说法,开角龙亚科约在8000至9000万年前起源于拉腊米迪亚,并于7700万年前分散在这片陆地的大部分区域。此时犹他州和科罗拉多地区出现一道屏障,阻止动物在拉腊米迪亚南北部之间扩散,并导致7700至7580万年前南北开角龙亚科(可能还有其它演化支)通过按纬度排列的孤立特有种中心内的地理隔离而独立进化。约7570万年前隔离消失后,前先一直困在拉腊米迪亚南部的华丽角龙谱系(以迷乱角龙谱系为代表)向北扩散,并进化出后来所有的开角龙亚科,如准角龙和三角龙。[2]
尽管坎帕阶后期拉腊米迪亚古陆上的恐龙比多数现代大型动物都要巨大(进食需求量增长,导致物种分布范围扩大),但桑普森和同事发现其分布范围可能较小,然而这片大陆上的恐龙物种多样性较高,因此该现象更令人困惑。这些恐龙显然居住在至少两块半隔离区域,但没有证据表明地区间存在扩散障碍,而且当时的气温梯度也比现在更小。当地恐龙似乎对环境的纬度性成带现象(Latitudinal zonation)非常敏感(可能由生理机能较低或环境生产力较高所致),该现象可能持续了至少150万年。其它假说包括孤立的“恐龙省”被混合动物区系分隔,或整块大陆上存在一个无特有种带的连续渐变群。阻止扩散的可能物理障碍包括存在东西向未知山脉及主要河流系统或西部内陆海道在平原地区泛洪(可暂时毁灭拉腊米迪亚中部的低海拔栖息地)。桑普森及其同事认为,当时很可能存在阻止扩散的古气候或古环境障碍(该假说得到拉腊米迪亚南北花粉类型不同的支持),但指出需要更多证据以调查拉腊米迪亚动物区系间的地理隔离。[2]
古生物学家泰瑞·盖茨(Terry A. Gates)和同事于2012年提出,坎帕阶北美恐龙的生物多样性增加是由西部内陆盆地的造山运动(导致大陆板块交汇处的地壳变化)所致,包括形成山脉以隔离角龙科和鸭嘴龙科,并导致其多样化的塞维尔造山带与拉腊米迪亚板块构造的早期融合。他们认为,华丽角龙和迷乱角龙从包括准角龙及三角龙在内的演化支中分化出来是由隔离或扩散事件所致。[44]坎贝尔2014年得出结论称迷乱角龙和开角龙的关系近于华丽角龙,认为迷乱角龙起源自拉腊米迪亚北部,并质疑桑普森和同事称其从南方迁徙至此的假说(迷乱角龙意即“流浪有角面孔”,该命名可能有用词不当之嫌)。[22][1]

2014年,朗里奇将颅骨CMN 8801归入华丽角龙未定种,并命名五角龙新种北方五角龙,两者皆来自阿尔伯塔省的恐龙公园组。朗里奇以此反驳划分南北“恐龙省”的想法,因为两者现在于北美东南及北方皆有分布。鉴于南北方动物确实不同,他同意坎帕阶存在明显的特有种分布,但发现恐龙谱系可以远距离扩散且不受地理障碍、气候或植物群的限制,因此该现象是已有恐龙种群(已适应当地环境)通过竞争淘汰后来者(迁徙至此的恐龙)的结果。[6]2016年,坎贝尔及其同事将CMN 8801重新归入开角龙,并支持华丽角龙仅分布于拉腊米迪亚南部的假说。[16]同年,马龙和同事发现北方五角龙是个无效的疑名,并同意拉腊米迪亚南北之间存在扩散障碍,指出华丽角龙和蒙大拿州北部钉盾龙之间的密切关系显示,两者之间存在某种短暂的地理联系。[27]
2016年,古生物学家斯宾塞·卢卡斯(Spencer G. Lucas)和同事批评拉腊米迪亚恐龙南北区域性分布的假说,并发现非恐龙脊椎动物的特有种分布现象极其罕见,况且主要通过几个开角龙亚科分类单元来鉴别该现象的做法存在问题。他们反驳了恐龙特有性及地方性分布的想法,理由包括:采样偏差存在问题(坎帕阶恐龙多样化的假象源自当时密度更高的化石记录)、缺乏可划分“恐龙省”的地形屏障(盖茨和同事可能误解了造山运动对拉腊米亚的地形影响)、缺乏显著的气候或植被差异、对该观点的理解涉及分类学决策、大多数化石群的穿时性(地质年龄差异)妨碍其在生物地理学分析中的使用以及支持该观点的人的结论不一致以致于违背论点。例如,他们指出华丽角龙和犹他角龙并非迷乱角龙(较年轻)的同时期物种,且三者的生存年代皆早于五角龙,与桑普森和同事的结论相反。卢卡斯和同事承认存在无法通过抽样偏差来解释的特有分布现象,但同意朗里奇的观点,即这些可能是由竞争或饮食特化所造成。[45]
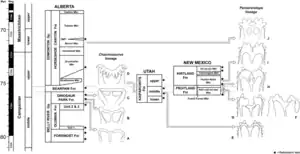
福勒2017年指出,桑普森和同事曾以凯帕罗维兹组的地质年龄证明华丽角龙和犹他角龙与北方其它地层中的开角龙亚科是同期物种,但年代校准与其它地层不同;重新校准到相同标准时,并未发现这些分类单元之间存在地层重叠。相反,凯帕罗维兹组仅在下层与恐龙公园组重叠,但重叠部分没有华丽角龙和犹他角龙化石,而零散遗骸表明两个地层在这部分存在相同分类单元。因此,福勒发现地层间恐龙分类单元的差异更可能是由采样不同地层而非生物地理隔离造成,而凯帕罗维兹组与果园组和科特兰组之间的差异可能是另一种解释。[37]虽然福勒和弗里德曼·福勒不支持恐龙存在特有种,但在2020年提出开角龙亚科从北到南有一种微妙且层次分明的地方分布。开角龙谱系在大陆北方更丰富,华丽角龙是其最南端的成员,而五角龙谱系在南方更丰富。由于中坎帕阶末期两个演化支的地理分布重叠,因此产生这两个谱系的物种形成事件可能是由7700万年前第一个开角龙谱系成员出现前的纬度差异造成。7600万年前,分隔两者的所谓“屏障”肯定已经消失,因为两者后来已经共存,而该分界线可能位于犹他州南部和蒙大拿州北部之间。[28]福勒和弗里德曼·福勒还发现,迷乱角龙生活在约7620万年前,而华丽角龙生活在约7690万年前,分布地层不存在重叠。[17]
参见
- 角龙类研究历史
参考资料
| 維基物種上的相關:華麗角龍屬 |
- . University of Utah News. 2010 [July 3, 2019]. (原始内容存档于February 22, 2011).
- Sampson, S. D.; Loewen, M. A.; Farke, A. A.; Roberts, E. M.; Forster, C. A.; Smith, J. A.; Titus, A. L.; Stepanova, A. . PLoS ONE. 2010, 5 (9): e12292. Bibcode:2010PLoSO...512292S. PMC 2929175
 . PMID 20877459. doi:10.1371/journal.pone.0012292.
. PMID 20877459. doi:10.1371/journal.pone.0012292. - Loewen, M.; Farke, A. A.; Sampson, S. D.; Getty, M. A.; Lund, E. K.; O’Connor, P. M. . Titus, A.; Loewen, M. (编). . Bloomington: Indiana University Press. 2013: 488–503. ISBN 978-0-253-00883-1.
- Getty, M. A.; Loewen, M. A.; Roberts, E.; Titus, A. L.; Sampson, S. D., , Ryan, M. J.; Chinnery-Allgeier, B. J.; Eberth, D. A. (编), , Bloomington: Indiana University Press: 478–494, 2010, ISBN 978-0253353580
- Sampson, S. D. . Scientific American. 2012, 306 (3): 40–47. Bibcode:2012SciAm.306c..40S. ISSN 0036-8733. JSTOR 26014255. PMID 22375321. doi:10.1038/scientificamerican0312-40.
- Longrich, N. R. . Cretaceous Research. 2014, 51: 292–308. doi:10.1016/j.cretres.2014.06.011.
- Levitt, C. G. (学位论文). University of Utah. 2013 [June 25, 2019]. (原始内容存档于July 14, 2017).
- Dodson, P. . Canadian Journal of Earth Sciences. 2013, 50 (3): 294–305. Bibcode:2013CaJES..50..294D. doi:10.1139/cjes-2012-0085.
- Gonzales, R.; Siegler, K.; Dwyer, C. . NPR.org. 2017 [June 25, 2019]. (原始内容存档于July 5, 2019).
- Finnegan, M. . Los Angeles Times. 2017 [June 24, 2019]. (原始内容存档于June 25, 2019).
- Panciroli, E. . The Guardian. 2017 [June 25, 2019]. (原始内容存档于May 31, 2019).
- Wei-Haas, M. . Smithsonian Magazine. 2017 [June 24, 2019]. (原始内容存档于June 24, 2019).
- Gramling, C. . Science. 2017 [June 26, 2019]. doi:10.1126/science.aal1122. (原始内容存档于June 26, 2019).
- . NRDC. April 10, 2020 [April 19, 2020]. (原始内容存档于2021-04-02).
- Longrich, N. R. . F1000Research. 2015, 4: 1468. doi:10.12688/f1000research.7573.1 .
- Campbell, J. A.; Ryan, M. J.; Holmes, R. B.; Schröder-Adams, C. J.; Mihlbachler, M. C. . PLOS ONE. 2016, 11 (1): e0145805. Bibcode:2016PLoSO..1145805C. PMC 4699738 . PMID 26726769. doi:10.1371/journal.pone.0145805.
- Fowler, D. W.; Freedman Fowler, E. A. . PeerJ. 2020, 8: e9251. PMID 32547873. doi:10.7717/peerj.9251/supp-1 .
- Paul, G. S. 2. Princeton: Princeton University Press. 2016: 285–286, 296. ISBN 978-0-691-16766-4.
- Mallon, J. C.; Holmes, R.; Eberth, D. A.; Ryan, M. J.; Anderson, J. S. . Journal of Vertebrate Paleontology. 2011, 31 (5): 1047–1071. doi:10.1080/02724634.2011.601484.
- Wick, S. L.; Lehman, T. M. . Naturwissenschaften. 2013, 100 (7): 667–682. Bibcode:2013NW....100..667W. PMID 23728202. doi:10.1007/s00114-013-1063-0.
- Longrich, N. R. . Cretaceous Research. 2011, 32 (3): 264–276. doi:10.1016/j.cretres.2010.12.007.
- Campbell, J. A. (学位论文). Carleton University. 2014 [June 30, 2019]. doi:10.22215/etd/2014-10978 . (原始内容存档于June 30, 2019).
- Campbell, J. A.; Ryan, M. J.; Schröder-Adams, C. J.; Holmes, R. B.; Evans, D. C. . Vertebrate Anatomy Morphology Palaeontology. 2019, 7: 83–100. doi:10.18435/vamp29356 .
- Fowler, D. W.; Scanella, J.; Horner, J. R. . Journal of Vertebrate Paleontology, SVP Program and Abstracts Book: 111A. 2011.
- 亦发表作Fowler, D. W.; Scanella, J.; Horner, J. R. . Journal of Vertebrate Paleontology. 2011, 31 (Supplement 2): 111. doi:10.1080/02724634.2011.10635174.
- Brown, C. M.; Henderson, D. M. . Current Biology. 2015, 25 (12): 1641–1648. PMID 26051892. doi:10.1016/j.cub.2015.04.041 .
- Lehman, T. M.; Wick, S. L.; Barnes, K. R. . Journal of Systematic Palaeontology. 2016, 15 (8): 641–674. doi:10.1080/14772019.2016.1210683.
- Mallon, J. C.; Ott, C. J.; Larson, P. L.; Iuliano, E. M.; Evans, D. C.; Evans, A. R. . PLOS ONE. 2016, 11 (5): e0154218. Bibcode:2016PLoSO..1154218M. PMC 4871577 . PMID 27191389. doi:10.1371/journal.pone.0154218.
- Fowler, D. W.; Freedman Fowler, E. A. . PeerJ. 2020, 8: e9251. doi:10.7717/peerj.9251 .
- Hedrick, B. P.; Goldsmith, E.; Rivera-Sylva, H.; Fiorillo, A. R.; Tumarkin-Deratzian, A. R.; Dodson, P. . The Anatomical Record. 2019: 40. PMID 30773832. doi:10.1002/ar.24099 .
- Ridgwell, N. M. . Museum and Field Studies Graduate Theses & Dissertations (学位论文). 2017 [June 30, 2019]. (原始内容存档于June 30, 2019).
- Padian, K.; Horner, J. R. . Journal of Zoology. 2011, 283 (1): 3–17. doi:10.1111/j.1469-7998.2010.00719.x .
- Knell, R. J.; Sampson, S. (PDF). Journal of Zoology. 2011, 283 (1): 18–22 [June 25, 2019]. doi:10.1111/j.1469-7998.2010.00758.x . (原始内容存档 (PDF)于July 22, 2018).
- Hone, D. W. E.; Naish, D. . Journal of Zoology. 2013, 290 (3): 172–180. doi:10.1111/jzo.12035 .
- Knapp, A.; Knell, R. J.; Farke, A. A.; Loewen, M. A.; Hone, D. W. E. . Proceedings of the Royal Society B: Biological Sciences. 2018, 285 (1875): 20180312. PMC 5897650 . PMID 29563271. doi:10.1098/rspb.2018.0312.
- Raia, P.; Passaro, F.; Carotenuto, F.; Maiorino, L.; Piras, P.; Teresi, L.; Meiri, S.; Itescu, Y.; Novosolov, M.; Baiano, M. A.; Martinez, R.; Fortelius, M. . The American Naturalist. 2015, 186 (2): 165–175. PMID 26655146. doi:10.1086/682011. hdl:11336/55178 .
- Roberts, E. M.; Deino, A. L.; Chan, M. A. . Cretaceous Research. 2005, 26 (2): 307–318. doi:10.1016/j.cretres.2005.01.002.
- Fowler, D. W.; Wong, William O. . PLOS ONE. 2017, 12 (11): e0188426. Bibcode:2017PLoSO..1288426F. PMC 5699823 . PMID 29166406. doi:10.1371/journal.pone.0188426.
- Sampson, S. D.; Loewen, M. A.; Roberts, E. M.; Getty, M. A. . Titus, A.; Loewen, M. (编). . Bloomington: Indiana University Press. 2013: 599–622. ISBN 978-0-253-00883-1.
- Roberts, E. M.; Sampson, S. D.; Deino, A. L.; Bowring, S. A.; Buchwaldt, S. . Titus, A.; Loewen, M. (编). . Bloomington: Indiana University Press. 2013: 85–106. ISBN 978-0-253-00883-1.
- Titus, A. L.; Eaton, J. G.; Sertich, J. . Geology of the Intermountain West. 2016, 3: 229–291. doi:10.31711/giw.v3i0.10.
- Wiersma, J. P.; Irmis, R. B. . PeerJ. 2018, 6: 76. PMC 6063217 . PMID 30065856. doi:10.7717/peerj.5016.
- Oser, S. E. (学位论文). Boulder: Department of Museum and Field Studies, University of Colorado. 2018 [July 5, 2019]. (原始内容存档于July 5, 2019).
- Sampson, S. D.; Loewen, M. A., , Ryan, M. J.; Chinnery-Allgeier, B. J.; Eberth, D. A. (编), , Bloomington: Indiana University Press: 405–427, 2010, ISBN 978-0253353580
- Gates, T. A.; Prieto-Márquez, A.; Zanno, L. E.; Dodson, P. . PLoS ONE. 2012, 7 (8): e42135. Bibcode:2012PLoSO...742135G. PMC 3410882 . PMID 22876302. doi:10.1371/journal.pone.0042135.
- Lucas, S. G.; Sullivan, R. M.; Lichtig, A.; Dalman, S. . New Mexico Museum of Natural History and Science Bulletin. 2016, 71: 195–213.