植龍目
植龍目(学名:Phytosauria)是群大型半水生掠食動物,身長2到12公尺,繁盛於三疊紀晚期。植龍目屬於镶嵌踝类,只有一科副鱷科(Parasuchidae)。植龍目具有長口鼻部與厚重鱗甲,在體型、外表、以及生活方式等方面與現代鱷魚類似,這是一個平行演化或趨同演化的例子。植龍目的名稱 「Phytosauria」意為「植物蜥蜴」,容易被人誤解為植食性動物,但牠們用來猛咬的頜部顯示出牠們是掠食動物。但首次敘述植龍目的人,誤以為他所研究的標本是植食性動物。
| 植龙目 化石时期:三疊紀 | |
|---|---|
 | |
| 卡羅萊納狂齒鱷復原圖 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | Reptilia |
| 演化支: | 真 Eureptilia |
| 演化支: | 卢默龙类 Romeriida |
| 亚纲: | 双孔亚纲 Diapsida |
| 演化支: | 新双弓类 Neodiapsida |
| 演化支: | 蜥类 Sauria |
| 演化支: | 主龜龍類 Archelosauria |
| 下纲: | 主龍形下綱 Archosauromorpha |
| 演化支: | 鱷腳類 Crocopoda |
| 演化支: | 主龍型類 Archosauriformes |
| 演化支: | 真鱷腳類 Eucrocopoda |
| 演化支: | 镶嵌踝类 Crurotarsi |
| 演化支: | 主龍類 Archosauria |
| 目: | †植龙目 Phytosauria von Meyer, 1861 |
多年来,植龍目被认为是“鑲嵌踝類主龙”(伪鳄类旧称)的最基础分支,最近对早期植龍目进化关系的一些研究表明,植龍目在伪鳄类和鸟跖类分化之前进化,并且是主龙类的姐妹分类群。
植龍目的分佈廣泛,化石已在歐洲、北美洲(尤其是亞伯達省)、印度、摩洛哥、泰國、以及馬達加斯加等地發現。在早侏罗世岩层中发现了疑似植龍目的化石,可能将其生存时间范围延伸到侏罗纪。
分类
- ?箭齒龍 Belodon
- ?尖齒鱷 Centemodon
- 优雅龙属 Compsosaurus
- 滇东鳄属 Diandongosuchus
- 佛朗哥鳄属 Francosuchus
- 中喙龙 Mesorhinosaurus
- 植龙 Phytosaurus
- 副鳄科 Parasuchidae
早期發現
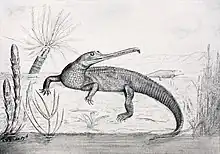
當第一個植龍目化石被發現時,並不清楚牠們是什麼動物。第一個被命名的種是植龍屬(Phytosaurus cylindricodon),意為「有圓柱形牙齒的植物蜥蜴」,由G. Jaeger在1828年命名,但他將頜部縫隙中的化石化泥土錯誤鑑定是牙齒。這個標本因保存不好而難以鑑識,而這個種名目前是無效名稱。在1861年,德國古生物學家克莉斯汀·艾瑞克·赫爾曼·汪邁爾(Christian Erich Hermann von Meyer)建立植龍目(Phytosauria),以最早被命名的植龍屬作為名稱來源。
第二個被命名種是箭齒龍(Belodon plieningeri),由汪邁爾與T. Plieninger在1844年所敘述、命名。在1875年,湯瑪斯·亨利·赫胥黎(Thomas Henry Huxley)將發現於印度的一個部分口鼻部化石命名為副鱷(Parasuchus hislopi),並加上植龍、箭齒龍,建立了副鱷目(Parasuchia),因為牠們有很大程度類似鱷魚。但当时因為這標本沒有足夠的鑑識特徵,所以副鱷被古喙龍(Paleorhinus)所取代。但是植龍科與副鱷科仍被不同科學家同時分別使用。植龍目(Phytosauria)是這些動物的標準名稱,名稱容易讓人誤解為植食性,但事實上牠們是肉食性動物。
與鱷魚的差異
儘管植龍目與鱷魚在外表與生活方式有許多相似處,仍有許多次要特徵可分辨這兩群動物。例如,植龍目的踝部結構比任何鱷魚的踝部結構還原始。植龍目也缺乏骨質次生顎,而鱷魚擁有次生顎,次生顎可讓鱷魚嘴部充滿水時仍保持呼吸。然而植龍目可能擁有肉質構成的次生顎,許多中生代鱷魚也被推測擁有肉質次生顎。最後,也是兩者最明顯的差別,植龍目的鼻孔位於眼睛附近或上方,而鱷魚的鼻孔位於口鼻部尾端。這些演化適應可讓牠們兩者的身體浸泡在水面下時,仍可以呼吸。
三種形態型

植龍目的頭顱骨有三種明顯的形態型(Morphotype),這些形態型與演化關係毫無關係,而與食性、習性有關聯。這些頭顱形態是以齒列特徵來分別;特別是頜部牙齒的差異與相似處。
Dolichorostral(長口鼻部):擁有細長口鼻部,以及佈滿大量的圓錐狀牙齒。牠們可能是魚食性,能夠捕抓快速、光滑的獵物,但無法順利追捕陸地動物。成員包括:古喙龍、狂齒鱷、以及迷鱷。過去一度認為古喙龍與迷鱷因為擁有同種適應演化,而屬於一個個別演化支(迷鱷科或迷鱷亞科),但現在迷鱷被認為與偽帕拉鱷有較近親緣關係。
Brachyrostral(短口鼻部):擁有巨大、寬廣的口鼻部,以及非常強壯的頭顱、頜部;前段牙齒類似犬齒狀,可緊握獵物,後段牙齒類似刀鋒狀,可將獵物切成大塊以利吞食(異齒型動物)。這些強壯的動物可將來到河邊飲水的陸地動物為食。成員包括:尼克羅龍、鑿齒鱷。
Altirostral(高口鼻部):為以上兩型的中間型。擁有不同齒列,但沒有短口鼻部型的極度發展。典型成員包括:狹喙龍(Angistorhinus)、偽帕拉鱷。牠們的食性極可能同時包括魚類、陸地動物。
現代鱷魚擁有類似的多樣形態型,例如寬廣口鼻部的短吻鱷,以及長口鼻部的食魚鱷。
植龍目有比鱷魚更好的裝甲保護,擁有厚重鱗甲(常與化石一起發現),腹部由密集排列的腹肋強化。
演化歷史與演化關係

植龍目首次出現於中三疊紀拉丁尼阶,從某種鑲嵌踝類演化而來。已知最早的植龍目已有完整外形與高度特化特徵,目前還沒有發現中間過渡物種。
已知最早的植龍目為原始、非特化的古喙龍,牠們分佈不廣泛。而稍微先進、較大的狹喙龍(Angistorhinus)出現在同一期間或稍晚。在卡尼階晚期,以上兩種動物被更特化的狂齒鱷、Leptosuchus、以及巨大的鑿齒鱷所取代。卡尼階-諾利階滅絕事件將以上物種滅絕了,諾利階早期出現了新屬,例如尼克羅龍與偽帕拉鱷,兩者都屬於更衍化的植龍目演化支,名為偽帕拉鱷亞科。在諾利階中期,出現了先進、特化、魚食性的迷鱷。迷鱷分佈廣泛,化石發現於北美洲西南部、義大利北部、以及泰國。最後,發現於北美洲西南部的大型雷東達龍(Redondasaurus),以及發現於歐洲、長口鼻部的Angistorhinopsis,生存於三疊紀的最後一期瑞提階。在美國東岸的瑞提階最晚期地層(紐瓦克超群)發現了植龍目的足跡。這顯示植龍目在三疊紀末前仍是成功的動物群,直到三疊紀-侏儸紀滅絕事件將植龍目滅亡。植龍目的滅絕比外表類似植龍的鱷魚出現,早了5000萬年;侏儸紀中早期的早期真正鱷魚,仍然體型小,且為完全陸生或完全海生。
參考資料
- Carroll, RL (1988), Vertebrate Paleontology and Evolution, WH Freeman & Co.
- Chatterjee, S (1978), A primitive parasuchid (phytosaur) reptile from the Upper Triassic Maleri Formation of India, Palaeontology 21: 83-127
- Hungerbühler A. (2002), The Late Triassic Phytosaur Mystriosuchus Westphali, With A Revision of the Genus. Palaeontology, March 2002, vol. 45, no. 2, pp. 377–418(42)
- Lucas, SG (1998), Global Triassic tetrapod biostratigraphy and biochronology. Paleogeog. Palaeoclimatol., Palaeoecol. 143: 347-384.
- Olsen, P.E., Kent, D.V., H.-D.Sues,, Koeberl, C., Huber, H., Montanari, E.C.Rainforth, A., Fowell, S.J., Szajna, M.J., and Hartline, B.W., (2002) Ascent of Dinosaurs Linked to an Iridium Anomaly at the Triassic-Jurassic Boundary. Science, vol. 296, p. 1305-1307.
外部連結
- 植龍目簡介(英文)
- 植龍目生活習性的初步研究(英文)
- 植龍目 - Palaeos網站
- 分類樹 - Mikko's Phylogeny網站
- 大型三疊紀動物群 - Chinle與紐華克生物群 (页面存档备份,存于)(英文)