南極甲龍屬
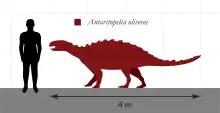
南极甲龙(属名:Antarctopelta,发音:/ænˌtɑːrktoʊˈpɛltə/ ann-TARK-toh-PEL-tə,意为“南极盾甲”)是甲龙类恐龙的一个属,生存于晚白垩世的南极,仅含一个已知物种奥氏南极甲龙(A. oliveroi)。它是种身长不超过4米(13英尺)的中型甲龙类,兼具两个不同科的特征,导致很难作出准确分类。单个已知化石标本于1986年在詹姆斯罗斯岛发现,是南极发现的第一件恐龙化石兼第二种正式命名的恐龙。
| 南极甲龙属 化石时期:晚白垩世, | |
|---|---|
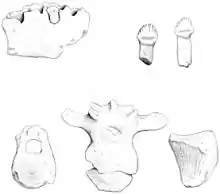 | |
| 正模标本的素描图 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | †鸟臀目 Ornithischia |
| 亚目: | †甲龙亚目 Ankylosauria |
| 演化支: | †副甲龍類 Parankylosauria |
| 属: | †南极甲龙属 Antarctopelta Salgado & Gasparini, 2006 |
| 模式種 | |
| †奥氏南极甲龙 Antarctopelta oliveroi Salgado & Gasparini, 2006 | |
发现与命名

正模标本是该物种的唯一已知标本,也是南极发现的首个恐龙化石,由三颗单个牙齿、部分下颌骨及另一颗处在原位的牙齿、其它颅骨碎片、椎骨(颈椎、背椎、臀部椎骨及尾椎)、部分肢骨(肩胛骨、髂骨及股骨)、脚趾骨骼(跖骨及趾骨)及大量甲片组成,约含完整骨骼的15%。标本由阿根廷地质学家爱德华多·奥利韦罗(Eduardo Olivero)和罗伯托·斯卡索(Roberto Scasso)于1986年1月在南极半岛外的詹姆斯罗斯岛首次发现,但冻土及恶劣天气导致挖掘工作花费近十年才完成。这些材料是在几个野外季期间从6 m2(65 sq ft)的区域内收集,但被认为属于同一个体。多数骨骼保存状况较差,因为许多离地表最近的化石因多年冻融风化而碎裂。[1]
尽管这些材料已发现数十年,并先后在三份不同出版物上发表,但直到2006年才由阿根廷古生物学家莱昂纳多·萨尔加多(Leonardo Salgado)和佐兰·加斯帕里尼(Zulma Gasparini)命名为奥氏南极甲龙(Antarctopelta oliveroi),因此它虽然是南极发现的第一种恐龙,却成为继1993年命名的冰冠龙之后命名的第二种南极恐龙。[2]属名意指该恐龙发现于南极大陆且拥有装甲,其中Antarctica一词源自希腊语αντ/ant-(与……相反)及αρκτος/arktos(熊,指朝向北方的大熊星座),[1]而希腊语πελτη/pelte(盾)是命名甲龙类(如雪松甲龙和蜥结龙)的常用词。种名纪念正模标本发现者爱德华多·奥利韦罗,他在出版物上首次提及南极甲龙,并在南极工作数十年之久。[1]
早期研究显示詹姆斯罗斯岛的甲龙类是只幼龙,[3][4]但近期研究表明椎骨不同部位已完全融合,而幼龙神经弓和椎体之间应该有可见骨缝。对几块骨骼的初步组织学分析也表明其存在一定程度的骨重塑,所以不是新形成的骨骼。[1]
描述

与其他甲龙类一样,奥氏南极甲龙是种矮壮的四足食植动物,全身被嵌入皮肤的装甲所保护。虽然尚未发现完整骨骼,但估计该物种从鼻部至尾末端的最大长度为4米(13英尺)。[1]2010年,格雷戈里·保罗(Gregory Paul)给出6米(20英尺)长、350(770磅)重的更高值。[5]目前对颅骨所知甚少,但所有已知颅骨碎片皆高度骨化以用作防御,尤其是一块鉴定为眶上骨的骨髂,上面有一支短刺,可能向外突出至眼睛上方。牙齿呈叶片状且不对称,多数小齿皆位于最靠近鼻尖的边缘。牙齿亦成比例地大于其它甲龙类,最大直径为10 mm(0.39英寸)。[1]相比之下,北美的包头龙体型更大,身长6至7米(20至23英尺),牙齿平均直径仅为7.5 mm(0.30英寸)。[6]
南极甲龙遗骸中已发现六种不同类型的皮内成骨,但很少与骨骼相连,因此在体表的排列方式很大程度上皆为推测。其中包括一支大型尖刺的基部。一些扁平矩形骨板类似结节龙科粗齿埃德蒙顿甲龙保护颈部的骨板。还发现一些大型圆形骨板,与较小多边形结节连接,可能像蜥结龙般在臀部形成盾甲。另一种皮内成骨呈椭圆形,并长有从中央向下延伸的龙骨。第五种皮内成骨附着在肋骨上,表明它们沿动物肋腹成行排列,在甲龙类中是种很典型的排列模式。最后一种主要由小型骨质结节组成,一般称为“小骨”,可能遍布全身。还发现几根肋骨上附着小骨。[1]
发现了一些尾椎。虽然尾部末端没有保存下来,但发现部分较小椎骨生前可能长于尾部末端,并与上下侧的骨化肌腱连接。甲龙科的肌腱有助于加固尾末端以支撑大型骨质尾槌。然而大型皮内成骨及特化尾椎表明,南极甲龙实际上可能长有锯剑般的扁平叶状尾部结构,类似于近亲覆尾龙。[7]
分类
南极甲龙与结节龙科存在几个共同特征,主要位于牙齿和装甲上,而疑似存在尾槌的尾部与甲龙科更相似。这种嵌合特征导致难以将其归入任何一科。在进行系统发育分析之前被划入甲龙类地位未定属。[1]后来汤普森等人2011年的系统发育分析显示,该属是已知最原始的结节龙科。[8]然而,2021一项研究发现南极甲龙属于南半球甲龙类一个独特的基干谱系――副甲龙类。[7]
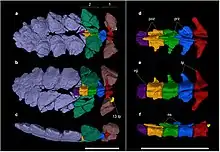
以下分支图取自西吉奥·梭托-阿库尼亚和同事2021研究中的系统发育分析:[7]
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
古生态学
正模标本骨骼收集于南极雪丘島組伽马段(Gamma Member)底部约90米(300英尺)处。[9]该层段沉积于浅海环境中,还保存着鲨鱼牙齿、沧龙类的南极拉库玛龙、菊石、双壳类及腹足类等海洋生物化石。菊石等标准化石显示这些岩石沉积于大约7400至7000万年前的晚白垩世坎帕阶晚期。[3]尽管南极甲龙发现于海洋沉积物中,但与所有甲龙类一样生活在陆地上。海洋沉积物中亦发现其它甲龙类,可能是尸体被冲入大海的结果。[10][11][12]
尽管白垩纪的南极洲位于南极区,但这段时期的地球气候更为温暖,因此该大陆不会结冰。像南极甲龙这样的动物可能生活在针叶林甚至落叶阔叶林中。尽管气温较高,但在冬天黑暗仍然会降临,就像今天的高纬度地区一样。[13]包括詹姆斯罗斯岛在内的南极半岛在此期间一直与南美相连,使两洲之间的动物群得以交流。事实上,近期发现的智利副甲龙类覆尾龙表明这些恐龙也居住在南美洲。[14]
参见
- 甲龙类研究历史
- 南极恐龙
参考资料
- Salgado, L.; Gasparini, Z. (PDF). Geodiversitas. 2006, 28 (1): 119–135 [2022-05-30]. (原始内容 (PDF)存档于2022-01-28).
- Hammer, William R.; Hickerson, William J. . Science. 1994-05-06, 264 (5160): 828–830 [2022-05-30]. Bibcode:1994Sci...264..828H. ISSN 0036-8075. S2CID 38933265. doi:10.1126/science.264.5160.828. (原始内容存档于2021-12-24) (英语).
- Olivero, E.; Gasparini, Z.; Rinaldi, C.; Scasso, R. . Thomson, M.R.A.; Crame, J.A.; Thomson, J.W. (编). . Cambridge: Cambridge University Press: 617–622. 1991.
- Gasparini, Z.; Pereda-Suberbiola, X.; Molnar, R.E. . Memoirs of the Queensland Museum. 1996, 39: 583–594.
- Paul, Gregory S. . New Jersey: Princeton University Press. 2010: 230.
- Vickaryous, M.K.; Maryanska, T.; Weishampel, D.B. . Weishampel, D.B.; Dodson, P.; Osmólska, H. (编). Second. Berkeley: University of California Press: 363–392. 2004.
- Soto-Acuña, Sergio; Vargas, Alexander O.; Kaluza, Jonatan; Leppe, Marcelo A.; Botelho, Joao F.; Palma-Liberona, José; Simon-Gutstein, Carolina; Fernández, Roy A.; Ortiz, Héctor; Milla, Verónica; Aravena, Bárbara; Manríquez, Leslie M. E.; Alarcón-Muñoz, Jhonatan; Pino, Juan Pablo; Trevisan, Cristine; Mansilla, Héctor; Hinojosa, Luis Felipe; Muñoz-Walther, Vicente; Rubilar-Rogers, David. . Nature. 9 December 2021, 600 (7888): 259–263. Bibcode:2021Natur.600..259S. PMID 34853468. S2CID 244799975. doi:10.1038/s41586-021-04147-1.
- Richard S. Thompson; Jolyon C. Parish; Susannah C. R. Maidment; Paul M. Barrett. . Journal of Systematic Palaeontology. 2011,. in press (2): 1. S2CID 86002282. doi:10.1080/14772019.2011.569091.
- Olivero, E.B. . Cretaceous Research. 2012, 34: 348–366. doi:10.1016/j.cretres.2011.11.015.
- Horner, J.R. . Journal of Paleontology. 1979, 53 (3): 566–577.
- Carpenter, K.; Dilkes, D.; Weishampel, D.B. . Journal of Vertebrate Paleontology. 1995, 15 (2): 275–297. doi:10.1080/02724634.1995.10011230.
- Lee Y.-N. . Journal of Vertebrate Paleontology. 1996, 16 (2): 232–245. doi:10.1080/02724634.1996.10011311.
- Huber, B.T. . Science. 1998, 282 (5397): 2199–2200. S2CID 129523803. doi:10.1126/science.282.5397.2199.
- . Nature. 1 December 2021: d41586–021–03572–6. PMID 34853419. S2CID 244830406. doi:10.1038/d41586-021-03572-6.