双生子研究
双生子研究(英語:)展现个体中环境与遗传分别造成的绝对、相对影响。双生子研究是行为遗传学研究的重要工具。行为遗传学研究还使用兄弟姐妹关系、领养关系、家族史等等信息。
双胞胎具有非常高的研究价值,用来研究不同家庭环境(不同对双胞胎所拥有的)与不同基因组成对性状造成的影响:同卵双生双胞胎有接近 100% 一样的基因,也就是双胞胎两者大部分不同(包括身高、无聊易感性、智能、抑郁程度等等)都是由双胞胎其中一位有而另一位没有的经验造成的。异卵双生双胞胎仅有 50% 的基因是共有的。双胞胎两者由于在相同时间、地点出生,拥有非常相似的环境因素(例如:子宫环境、 父母教养方式、教育、家庭富有程度、文化等等)。同卵双生双胞胎中其中一位有而另一位没有的性状为环境因素研究提供了有力的数据。
古典双生子研究将同卵双胞胎的相似性与异卵双胞胎的相似性进行比较,如果同卵双胞胎比异卵双胞胎更相似(在大部分性状都是如此),这表示基因在这些性状占有比较重要的角色。透过比较上百对双胞胎的家庭状况,研究人员得以更理解遗传因素、共享环境、独特环境分别在塑造人类行为上的重要性。
近代双子研究显示接近所有性状都具有遗传差异带来的部分影响,有些特征受到遗传比较强的影响(例如:身高),有些受到遗传中等程度的影响(例如:性格特质),而有些有复杂的遗传关系:在自闭症中不同的基因影响性状的不同部分。
历史
自从人类文明开始以来,学者就一直对双胞胎感兴趣。希波克拉底(公元前五世纪)将双胞胎共有的疾病归咎于双胞胎共有的物质条件而波希多尼(公元前一世纪)将共有的疾病归咎于双胞胎共有的星象条件[1]。后来弗朗西斯·高尔顿开创了透过双胞胎进行基因、环境对人类发育、行为影响的研究,然而高尔顿并没有发现同卵、异卵双胞胎之间的差异[2]。
这个差异在爱德华·桑代克(1905)为五十对双胞胎进行第一个心理学双生子测试的时候仍尚未被意识到,这个研究透过比较 9-10 岁、13-14 岁的双胞胎与一般手足的差异试图证明家庭对行为的影响随着年龄逐渐减弱。桑代克从他的数据不正确的推论出双胞胎只有一种类型。最早了解这个差异的可能是德国的遗传学家赫尔曼·沃纳·西门子在 1924 年的研究[3]。
方法
双生子设计的说服力来自于这一事实,即所有双胞胎分为单合子: 双胞胎由同一受精卵发育而来,因而所有等位基因相同,称为同卵双胞胎;及双合子 : 双胞胎分别由不同受精卵发育而来,因而平均只有50%的等位基因相同,该水平与非双胞胎的兄弟姐妹一致。这些在遗传相似性方面的已知区别,以及对同卵或异卵双胞胎所在环境相同的可验证的假设[4],共同构成了双胞胎设计的基础,以探索遗传多样性及环境多样性对某一形状的影响。[5][6]
双生子研究的逻辑基础只需很少的数学基础就能理解,包括对相关性的理解,以及对变异的概念的理解。
与所有的行为遗传学研究类似,传统的双生子研究从评估一个大群体中的行为多样性开始(遗传学家称为性状),并尽力估计其受以下因素影响的程度:
- 遗传效应(遗传度);
- 共同处在的环境 – 共同作用在双胞胎身上的环境因素,以同一方式影响两者;
- 各异的环境因素 – 不共同作用于双胞胎的环境因素。
作为代表,以上三个因素分别被简写为A (additive genetics,附加的遗传因素)、C (common environment,共同的环境因素) 和 E (unique environment, 各异的环境因素);因此三者的缩写连起来就是"ACE"。另外,估计非附加遗传因素所造成的影响也是可以的(由于全称是dominance故一般被标注为D(ADE模型);下文有更多关于复杂双生子设计的介绍)。
ACE模型揭示了对于影响某一性状的因素来说,可遗传的多样性所占比例是多大,以及共同环境因素以及各异环境因素所占比例是多少。该研究是通过SEM系统例如OpenMx来完成的,然而双生子设计的核心逻辑是不变的,如下所述:
在同一个家庭里长大的同卵双胞胎,其基因完全相同,环境完全相同,所有各异环境因素都是随机的。这种同卵双胞胎可以为我们提供A+C的参数估计。异卵双胞胎的C也是相同的,但彼此只有50%的基因是相同的: 因而这种异卵双胞胎可以直接用来估计½A+C。假设r是correlation相关系数,那么rmz和rdz就分别是同卵双胞胎和异卵双胞胎的性状相关系数。对于任一形状来说,有:
- rmz = A + C
- rdz = ½A + C
因而A等于同卵双胞胎和异卵双胞胎相关系数差异的两倍: 基因相加效应(Falconer's formula)。C就能通过同卵双胞胎相关系数减去A得到。同卵双胞胎的差异只由各异环境因素造成,因而随机(各异)因素E 等于1 − rmz。(Jinks & Fulker, 1970; Plomin, DeFries, McClearn, & McGuffin, 2001).
再次声明,以上两个加和式子可供我们解出A,C,E的值。由于同卵和异卵相关系数的差异是完全由只有一半的遗传相似性造成的,所以基因相加效应A就是相关系数差值的两倍:
- A = 2 (rmz − rdz)
由于同卵双胞胎的相关系数反映的是A和C的影响,因而E可以由1减去此两者而得到:
- E = 1 − rmz
最终, C可以由此得到:
- C = rmz − A
现代模型
从20世纪70年代开始,研究逐渐转向对遗传问题进行 建模,环境因素的影响通过最大似然模型得到(Martin & Eaves, 1977)。当计算变得越加得复杂,该方法表现出了许多优势,因而成为现在研究中的常用手段。
一个作为例子的结构模型展示如下( 为了估计身高的遗传度 ),其以丹麦男性数据为基础 (一个有代表性的群体中的8个独立多国队列)[7]
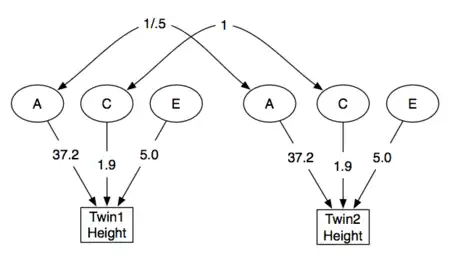 |
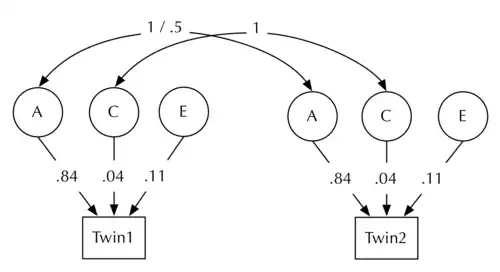 |
左边的模型展示了身高的原始多样性,其在用于得到基因和环境的绝对效应时很有用,即将效应量化成自然单位,比如研究身高改变时得到毫米级的指标。有时标准化参数是很有用的,这样各个参数可以直接作为总和的一部分而不用换算。由于我们已经把多样性分解为A,C,E了,所以多样性的总和直接就是A+C+E。我们可以将各个参数换算成其在总和中所占的比例,比标准化的A = A/(A+C+E):
模型比较
建模最基本的优势在于其能够对各个模型进行明确比较:比起传统的简单地对各个参数返回数值,建模可以计算参数的置信区间,但关键的是它能够去掉或添加路径,并通过统计学方法(如 AIC)检测其效果。举个例子,想要测试模型对于家庭或共同环境因素对性状的影响效应的话,我们可以用AE模型和ACE模型进行比较;就以上图的身高为例: C (共同环境因素) 被去掉的话会导致模型拟合程度的显著下降吗?另外,对于每一个路径来说,置信区间也能够被计算得到。
多群体和多变量模型
建模也包括多变量建模,其在用于解决明显是多变量之间的遗传关系的问题时非常有用。比如:智商和长期记忆之间有共同的基因吗?有共同的环境因素影响吗?多变量模型的优势还包括处理区间、阈值和连续数据,在数据包含缺失数值时保留全部数据,将测得变量与潜在模型进行整合,比如测得的环境变量,或者测得的分子遗传标记如SNPs。另外,建模能够避免把变量数值标准化,而是直接带入其原始值。
多变量和多时间波研究现在已成为了标准,其包含测得的环境变量和对潜在可能有因果关系的性状测量。其模型包括扩展的双生子设计,[8][9] 单一模型,[10] 以及生长曲线模型。[11]
SEM 系统(比如OpenMx[12] 以及其他适用于标准化数据及多群体的应用)使得这项新技术能够被有一定技能的人所使用。
对环境建模: 单合子不一致设计
由于同卵双胞胎的基因和家庭环境因素完全一样,其差异全部被归结于E: 各异环境因素。研究者可以由这一信息通过有力的方式来理解环境,包括流行病学中致病因素的检测,其在其他方法中会被其他因素(如基因-环境协方差,逆因果关系和混杂因素)混淆。 关于单合子不一致设计的效果,有一个例子如下左图所示。双胞胎中在性状1中得分更高的那个,在性状2中同样得分更高。这是因为性状1的“剂量”会导致性状2的增加。当然,性状2也会影响性状1。解出这两个概率需要一个不同的设计( 例子见诸下文 )。零假设是二者并无因果关系。
 “单合子不一致模型效应的例子” |
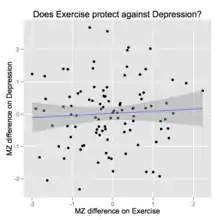 “MZ不一致测试的假设:运动能够保护我们免得抑郁症” |
举个例子,关于抑郁症和运动之间可见的联系( 见右上图 )。对于抑郁人群有报告称其存在运动较少的情况。因而我们或许会假设其存在因果关系:给抑郁症患者一定“剂量”的运动能够帮助他们精神振奋并缓解抑郁。下一幅图展示了对二者进行经验上的检验所得到的结果:空。[13]
纵向不一致设计
如下图所示,该设计可以被拓展到多次测量,因而模型就能反映出更加多元的信息。这被称为交叉滞后模型(对多性状进行多次测量)。[14]
在纵向不一致模型中,同卵双胞胎之间的差异能够被用于研究在时间点1时性状之间的差异(路径1),然后检测各个明确假设,如一个个体与另一个体相比在性状1上的数值盈余会导致其同一性状在未来的额外增量(路径B和E), 或者另一方面, 也是很重要的一方面, 对其他性状的额外增亮(路径C和D)。 在本例中,关于抑郁症患者的运动量低于人群平均水平,其二者的关联是有因果关系的假设,能够被成功验证。如果运动能够缓解抑郁症,那么路径D应该是显著的,即双胞胎中运动量更大的那个,抑郁程度应该更轻。

假设
从上面的模型中可以看出,双生子研究的主要假设是相同的环境。这个假设已经直接测试。特殊情况是,父母相信他们是异卵双胞胎,但事实上,他们基因完全相同。一系列心理特征的研究表明,这些孩子仍然像同卵双胞胎。<ref>Kendler, K.S., Neale, M.C., Kessler, R.C., Heath, A.C. and Eaves, L.J., 1993. Panic disorder in women: A population-based twin study. Psychological Medicine 23, pp. 397–406</ref> 分子遗传学的遗传力估算法提供了一些证据表明,双生子研究的经典设计——相同环境假设的可能是有效的,实实在在的,稳固的,合理的。
相似性测定:双生子研究假设的直接测试
Visscher等[15] 报道了一种针对双生子研究的强大的测试技术。他们利用了这一事实——兄弟姐妹平均有50%的基因相同,而对于特定的兄弟姐妹同胞对,相同基因数在这个数字附近,也就是说本质上是在家庭中创建一个连续的遗传相似性或“双胞胎”(填补普通同胞与双胞胎之间的空隙)。基于直接估计基因相似性来估计遗传力,为该方法的假设提供支持。
性别差异
两性之间的遗传因素可能不同,包括基因表达和基因环境相互作用。异性异卵双胞胎是在阐明这些效应中有重要价值。 一个极端的例子是,一个基因可能只在一种性别中表达(性别的定性限制)。通常,基因的影响可能受个人的性别的影响。基因可能导致男性体重改变100克,但也许在雌性中导致体重改变150克——基因的数量效应。一些环境可能影响基因发挥功能而环境是通过性别来产生影响。例如如果把女性排除在选举之外,影响投票行为的基因在女性中不发挥作用。更为普遍的是,性别差异测试的逻辑可以扩展到任意定义的子群的个体。在这样的情况下,同性和异性异卵双胞胎的相关性会不同,而与性别差异的影响无关。 由于这个原因,区分三种类型的异卵双胞胎是需要的。标准的分析流程将涉及测试性别限制通过拟合为五组——男性同卵双胞胎,女性同卵双胞胎,男性异卵双胞胎,女性异卵双胞胎,异性异卵双胞胎。双胞胎建模从而不仅是测试变量(如性别)的因果模型的相关关系。
基因环境相互作用
基因影响可能会依赖于环境。这种相互作用被称为“基因-环境相互作用”,在一个基因的某种基因型在不同的环境产生的效应不同。一个简单的例子是一个基因与环境的影响协同:在高营养环境中可能让身高增加1英寸,但在低营养环境中只让身高增加0.5英寸。对于不同的基因型,个体对不同环境的反应程度也不同。 通常研究者感兴趣于在不同条件下遗传效应的变化:在某种环境中,等位基因可以对表型产生较大的影响(如上)。 第二个作用是“基因环境相关性”,也就是说在特定的等位基因往往处于特定的环境。如果一个基因导致父母喜欢阅读,由于基因环境的相关性,继承这个等位基因的孩子更可能在有书的家庭长大:一个或两个父母有这个等位基因,因此会在家庭积累书籍并且传递阅读的等位基因。这种效应可以直接通过测量环境关联性(在本例中为在家里的书籍)来测试。 通常环境似乎在个体的最早期中作用最大,随着[义务教育]的开始作用迅速减少。这种现象可以在阅读[16]以及智力[17]观察到。这是基因-年龄效应的一个例子,并允许考量的由父母带来的(随着时间,作用慢慢减弱)基因-环境相关性和个体主动选择特定环境带来的基因-环境相关性。[18]
反应基线
研究植物或动物允许实验随机组合基因型和环境的来衡量各种变量的影响。相比之下,人类研究通常只能选择观察。[19][20] 这可能表明,反应基线难以评估。[21][22]
在其他领域如经济学和流行病学,几个设计开发利用能够利用不同的基因相似度,重复暴露,并测量暴露于环境(如儿童的社会阶级、家庭混乱,教育质量、营养、毒素等)来对抗那些混淆的因素。经典的双生子研究设计内在吸引力是其避开了这些混淆因素。例如,同卵及异卵双胞胎环境相同和基因的影响并不受到混淆。[6] 双胞胎的研究从而在一定程度上试图利用家庭成员基因的随机组合来帮助理解这些相关性。
而双生子研究只告诉我们在观察范围内的环境中基因和家庭如何影响行为,基因和环境将共变的警告,比起过去(不知道基因和环境的不同的角色),这是一个相当大的进步。[23] 因此双生子研究经常被用于至少控制观察变量中的一个部分的手段:例如把先前假定为家庭环境的因素划分到共同环境因素和附加遗传因素里面,这要通过完全相同或部分相同的双胞胎基因组实验来完成。[23]
没有单一的设计可以解决所有的问题。经典的双生子研究设计有额外可用的信息。双生子收养的研究设计是一种自然实验(将相同的基因型的个体置于不同的环境中)。[24]关联分析,如[25]允许直接研究等位基因的效应。等位基因的孟德尔随机化还提供了研究随机等位基因的影响对他们的相关环境和其他基因的机会,例如[26] 允许对等位基因效应的直接研究。等位基因的孟德尔随机化也给我们提供了研究等位基因在随机情况下效应的机会,其能考虑到与等位基因关联的环境和其他基因。例如[26]
拓展双生子研究设计,更多复杂的遗传学模型
基本的或经典的双生子研究设计只包含同卵和异卵双胞胎在其生物学家庭中成长情况。这只是所有可能的遗传和环境的关系的一个子集。因此,可以说,基于双生子研究的遗传估计只是理解的行为遗传学的第一步。 变量(双生子研究分为遗传相加性,共享环境,不共享环境)是完整研究基因-环境共变性,基因-环境相关性,其他对行为的非加和性的效应的一次尝试。分子遗传学的革命提供了更有效的描述基因组的工具,许多研究人员使用分子遗传学来直接评估等位基因和环境对表型的影响。 双生子研究设计的局限性首先是它不提供同时考虑共享环境和非加和基因效应的途径。这种限制可以通过添加额外的兄弟姐妹到设计中来解决。 第二个限制是基因-环境相关性不是作为确定无疑的效应来观察。解决这个限制需要整合收养模型,或采用双生子的子女研究设计,来评估相同基因时家庭的影响。
连续变量和有序变量
虽然一致性研究比较特征的存在或不存在于每一个双胞胎,相关研究比较在双胞胎不断变化表型的一致性。
批评
双生子方法一直以来受到来自统计遗传学、统计学及心理学界的批评,这些批评认为通过这种方法得到的结论是模糊或没有意义的。这些批评的核心要点和他们的陈述如下。
统计方法的批评
据称,双生子研究是没有统计学基础的。这种统计学批评认为大部分双生子研究的遗传力评估依靠在未经验证、有限制的假设之上,如果这样,那该研究的数据不能被严格信任。
例如,Peter Schonemann 就对20世纪70年代发展出的遗传力评估方法提出异议。他认为双生子研究中的遗传评估可能反映出别的因素而非共享基因本身。 使用发表在 Loehlin and Nichols (1976),[27] 的统计学模型对问题“你会擦背吗?”进行分析显示:该行为对0.92的男性存在遗传性,对0.21的女性存在遗传性,对问题“你会在天黑后佩戴太阳眼镜吗?”则对130%的男性存在遗传性,对103%的女性存在遗传性。[28][29]
对统计方法的批评的回应
在没有计算机以前,统计人员被迫使用有已知的限制,但计算易进行的方法。在20世纪80年代这些不精确的统计方法已经被丢弃:基于结构方程模型的现代双生子方法已经不受上方提到的因数学上不可能的影响。[30] 更重要的是,这些新的方法允许显性地测试各种通路上不同角色带来的复杂效应。[23]
取样:双胞胎作为代表性人群
双生子研究的结果不能认为是派生出他们的人群的研究结果。因此,明白这是基于特定样品或是有双胞胎本身特性的研究结果很重要。双胞胎不是人群中的随机样品,且他们的成长环境不同。在这个意义上说,他们不具有代表性。[31]
例如:异卵双胞胎的出生受很多因素影响。有些女性经常会在经期产生多于1个的卵子,因此她们更易怀有双胞胎。这个趋势在整个家族中都表现明显,通常父系与母系都是这样。年龄大于35岁的女性更易产生2个卵子。拥有3个及以上孩子的女性也更易拥有异卵双胞胎。人工诱导排卵及体外受精胚胎置入也可产生多胎和同卵双胞胎。[32][33][34][35][36][37]
对双胞胎代表性的回应
双胞胎与非双胞胎兄弟姐妹差别甚小。研究表明双胞胎与非双胞胎兄弟姐妹各自的个性及智力评估非常相似。(如 Deary et al. 2006).
以分离双胞胎为代表的其他双胞胎
分离双胞胎中,同卵或异卵多因被领养而分开。这使得他们原来的家庭并不具有代表性,因为他们放弃抚养孩子,而他们去的新家庭也不具有代表性,因为他们经过儿童保护机构批准,且大部分家庭没有亲生孩子。这些参与研究的志愿者甚至不能代表分离双胞胎,因为并非所有分离双胞胎都同意参与双生子研究。[38][39]
10%的环境影响因素不能解释弗林效应
一些如Steven Pinker的心理学家使用的数字因无法解释弗林效应而遭到批评,这些数字是45%遗传,45%天命(指随机性,无宗教含义),10%环境,按这些数字估算,20世纪早些时候的平均IQ提升过大,甚至超过了今天。按定义,随机个体的几率不会随时间显示出任何趋势。[40][41]
检出与否的问题
在人们选择保密及因被歧视等原因而曾经选择保密的行为案例中,存在事实未检出的问题。假如环境对该行为没有影响,但由于扭曲的结果,则可能使得环境因素看起来起了作用。对于环境似乎不起作用的案例,要不就是行为及其顽固,要不就是在研究中存在致使研究科学上不可用的缺陷。即使环境确实有影响,但数据仍会出错。[42][43][44]
Physical few-character stereotypes causing behavioral correlations
Apparent correlation between heredity and behavior may be due to social stereotypes making people with certain physical characteristics (which may be hereditary physical characteristics) behave in certain ways, without any biological link between the genetic factor and the behavior. This factor has an especially high likelihood of being the case for behaviors that are culturally depicted in mass media as being more accepted by those who consider them to be biological than by those who consider them learned (e.g. sexuality and some psychiatric diagnoses) and for physical characteristics that are not culturally considered by mass media to be bigoted to associate with certain behaviors (e.g. not ethnic or racial). Such unequal environment biases may concentrate on a relatively small number of minor physical characteristics that are subject to stereotypes that are considered to be politically correct, with no statistically significant effect on overall ratings of physical similarity, let alone anywhere near being able to pass identical twins off as being fraternal.[45][46][47]
Terminology
Pairwise concordance
For a group of twins, pairwise concordance is defined as C/(C+D), where C is the number of concordant pairs and D is the number of discordant pairs.
For example, a group of 10 twins have been pre-selected to have one affected member (of the pair). During the course of the study four other previously non-affected members become affected, giving a pairwise concordance of 4/(4+6) or 4/10 or 40%.
Probandwise concordance
For a group of twins in which at least one member of each pair is affected, probandwise concordance is a measure of the proportion of twins who have the illness who have an affected twin and can be calculated with the formula of 2C/(2C+D), in which C is the number of concordant pairs and D is the number of discordant pairs.
For example, consider a group of 10 twins that have been pre-selected to have one affected member. During the course of the study, four other previously non-affected members become affected, giving a probandwise concordance of 8/(8+6) or 8/14 or 57%.
参考资料
- Cicero, De Divinatione, (On Divination), ii. 42
- Richard D. Rende, Robert Plomin, Steven G. Vandenberg. . Behavior Genetics. 1990-03-01, 20 (2): 277–285 [2018-04-02]. ISSN 0001-8244. doi:10.1007/bf01067795. (原始内容存档于2018-06-18) (英语).
- Siemens, Hermann Werner. . Berlin: Springer. 1924. OCLC 18362377.
- Propping, Peter; Bouchard, Thomas J. (编). . London: J. Wiley. 1993: 326. ISBN 0-471-94174-3.
- Cardon, Lon R.; Neale, Michael C. . Boston: Kluwer Academic Publishers. 1992. ISBN 0-7923-1874-9.
- Nicholas Martin, Dorret Boomsma, Geoffrey Machin. . Nature Genetics. 1997-12-01, 17 (4): 387–392 [2018-04-02]. doi:10.1038/ng1297-387 (英语).
- Karri Silventoinen, Sampo Sammalisto, Markus Perola, Dorret I. Boomsma, Belinda K. Cornes, Chayna Davis, Leo Dunkel, Marlies de Lange, Jennifer R. Harris, Jacob V. B. Hjelmborg, Michelle Luciano, Nicholas G. Martin, Jakob Mortensen, Lorenza Nisticò, Nancy L. Pedersen, Axel Skytthe, Tim D. Spector, Maria Antonietta Stazi, Gonneke Willemsen, Jaakko Kaprio. . Twin Research and Human Genetics. 2003-10, 6 (5): 399–408 [2018-04-02]. ISSN 2053-6003. doi:10.1375/136905203770326402. (原始内容存档于2018-06-06) (英语).
- Matthew C. Keller, Sarah E. Medland, Laramie E. Duncan. . Behavior Genetics. 2010-05-01, 40 (3): 377–393 [2018-04-02]. ISSN 0001-8244. doi:10.1007/s10519-009-9320-x. (原始内容存档于2018-06-14) (英语).
- William L. Coventry, Matthew C. Keller. . Twin Research and Human Genetics. 2005-06, 8 (3): 214–223 [2018-04-02]. ISSN 1839-2628. doi:10.1375/1832427054253121. (原始内容存档于2018-06-03) (英语).
- Nathan A. Gillespie, David E. Evans, Margie M. Wright, Nicholas G. Martin. . Twin Research and Human Genetics. 2004-12, 7 (6): 637–648 [2018-04-02]. ISSN 2053-6003. doi:10.1375/1369052042663814. (原始内容存档于2018-06-02) (英语).
- Michael C. Neale, John J. McArdle. . Twin Research and Human Genetics. 2000-06, 3 (3): 165–177 [2018-04-02]. ISSN 2053-6003. doi:10.1375/136905200320565454. (原始内容存档于2018-06-02) (英语).
- Steven Boker, Michael Neale, Hermine Maes, Michael Wilde, Michael Spiegel, Timothy Brick, Jeffrey Spies, Ryne Estabrook, Sarah Kenny, Timothy Bates, Paras Mehta, John Fox. . Psychometrika. 2011-04-01, 76 (2): 306–317 [2018-04-02]. ISSN 0033-3123. doi:10.1007/s11336-010-9200-6. (原始内容存档于2018-06-13) (英语).
- M. H. De Moor, D. I. Boomsma, J. H. Stubbe, G. Willemsen and E. J. de Geus. (2008). Testing causality in the association between regular exercise and symptoms of anxiety and depression. Archives of general psychiatry, 65, 897-905. doi
- S. A. Burt, M. McGue and W. G. Iacono. (2009). Nonshared environmental mediation of the association between deviant peer affiliation and adolescent externalizing behaviors over time: results from a cross-lagged monozygotic twin differences design. Dev Psychol, 45, 1752-60. doi
- Peter M. Visscher, Sarah E. Medland, Manuel A. R. Ferreira, Katherine I. Morley, Gu Zhu, Belinda K. Cornes, Grant W. Montgomery, Nicholas G. Martin. . PLOS Genetics. 2006-03-24, 2 (3): e41 [2018-04-02]. ISSN 1553-7404. doi:10.1371/journal.pgen.0020041. (原始内容存档于2018-06-06) (英语).
- Brian Byrne, Sally Wadsworth, Robin Corley, Stefan Samuelsson, Peter Quain, John C. DeFries, Erik Willcutt, Richard K. Olson. . Scientific Studies of Reading: 219–235. [2018-04-02]. doi:10.1207/s1532799xssr0903_3. (原始内容存档于2021-06-20).
- Ian J Deary, Frank M Spinath, Timothy C Bates. . European Journal of Human Genetics. 2006-06, 14 (6): 690–700 [2018-04-02]. ISSN 1476-5438. doi:10.1038/sj.ejhg.5201588. (原始内容存档于2018-07-30) (英语).
- Robert Plomin, Denise Daniels. . Behavioral and Brain Sciences. 1987-03, 10 (1): 1–16 [2018-04-02]. ISSN 1469-1825. doi:10.1017/s0140525x00055941. (原始内容存档于2018-03-14) (英语).
revisited in: Robert Plomin, Denise Daniels. . International Journal of Epidemiology. 2011-06-01, 40 (3): 563–582 [2018-04-02]. ISSN 0300-5771. doi:10.1093/ije/dyq148. (原始内容存档于2018-03-18) (英语). - Oscar Kempthorne. . Genetica. 1997-03-01, 99 (2-3): 109–112 [2018-04-02]. ISSN 0016-6707. doi:10.1023/a:1018352202363. (原始内容存档于2018-06-04) (英语).
- Kenneth S. Kendler. . Archives of General Psychiatry. 1984-06-01, 41 (6) [2018-04-02]. ISSN 0003-990X. doi:10.1001/archpsyc.1984.01790170029004 (英语).
- Kamin, Leon J.; Rose, Steven R.; Lewontin, Richard C. . New York: Penguin Books. 1984. ISBN 0-14-022605-2.
- Richard J. Rose. . Science. 1982-02-19, 215 (4535): 959–960 [2018-04-02]. ISSN 0036-8075. doi:10.1126/science.215.4535.959. (原始内容存档于2018-06-04) (英语).
- M. C. Neale and H. H. Maes. (1996). Methodology for genetics studies of twins and families. Journal.
- S PETRILL. . Intelligence: 403–409. [2018-04-02]. doi:10.1016/j.intell.2004.05.001. (原始内容存档于2018-06-16).
- Colin D. Steer, George Davey Smith, Pauline M. Emmett, Joseph R. Hibbeln, Jean Golding. . PLOS ONE. 2010-07-13, 5 (7): e11570 [2018-04-02]. ISSN 1932-6203. doi:10.1371/journal.pone.0011570. (原始内容存档于2021-06-20) (英语).
- G. D. Smith. . Journal of the Royal Society of Medicine: 432–435. doi:10.1258/jrsm.100.9.432.
- Loehlin, John C.; Nichols, Robert. . Austin: University of Texas Press. 1976. ISBN 0-292-73003-9. JSTOR 2826060.
- Peter H. Schönemann. . Genetica. 1997-03-01, 99 (2-3): 97–108 [2018-04-02]. ISSN 0016-6707. doi:10.1023/a:1018358504373. (原始内容存档于2018-06-04) (英语).
- Schönemann, Peter H. . 1995 Meetings of the American Association for the Advancement of Sciences. 1995.
- M. C. Neale, S. M. Boker, G. Xie and H. H. Maes. (2002). Mx: Statistical Modelling. Journal.
- R. G. RECORD, THOMAS McKEOWN, J. H. EDWARDS. . Annals of Human Genetics. 1970-07-01, 34 (1): 11–20 [2018-04-02]. ISSN 1469-1809. doi:10.1111/j.1469-1809.1970.tb00215.x (英语).
- Woollett, Anne; Clegg, Averil. . London: Century. 1983: 127. ISBN 0-7126-0204-6. OCLC 12445470.
- S. L. Corson, R. P. Dickey, B. Gocial, F. R. Batzer, E. Eisenberg, L. Huppert, G. Maislin. . Fertility and Sterility. 1989-04, 51 (4): 644–650 [2019-05-25]. ISSN 0015-0282. PMID 2924931. (原始内容存档于2019-08-07).
- C. Derom, R. Derom, R. Vlietinck, H.Vanden Berghe, M. Thiery. . The Lancet: 1236–1238. [2018-04-02]. doi:10.1016/s0140-6736(87)92688-2. (原始内容存档于2018-06-16).
- R. G. Edwards, L. Mettler, D. E. Walters. . Journal of in Vitro Fertilization and Embryo Transfer. 1986-04-01, 3 (2): 114–117 [2018-04-02]. ISSN 0740-7769. doi:10.1007/bf01139357. (原始内容存档于2018-06-14) (英语).
- Leigh, Gillian. . London: Routledge and Kegan Paul. 1983. ISBN 9780710098887.
- Capron, Christiane; Vetta, Adrian R.; Duyme, Michel; Vetta, Atam. . Cahiers de Psychologie Cognitive/Current Psychology of Cognition. 1999, 18 (2): 115–160.
- Fatal Flaws in the Twin Study Paradigm: A Reply to Hatemi and Verhulst, Doron Shultziner 2013
- Twin Studies of Political Behavior: Untenable Assumptions?, Jon Beckwith and Corey A. Morris 2008
- Peters, Brad M. (2013). "Evolutionary psychology: Neglecting neurobiology in defining the mind". Theory & Psychology 23 (3)
- Ehrlich, Paul; Feldman, Marcus (2003). "Genes and Cultures: What Creates Our Behavioral Phenome?". Current Anthropology 44 (1)
- Critical Analysis: A Comparison of Critical Thinking Changes in Psychology and Philosophy Classes, Teaching of Psychology 2014 41: 28
- Association for Psychological Science: Why Science Is Not Necessarily Self-Correcting, John P. A. Ioannidis 2012
- How Black African and White British Women Perceive Depression and Help-Seeking: a Pilot Vignette Study, International Journal of Social Psychiatry March 2010
- (PDF). [2015-06-29]. (原始内容 (PDF)存档于2012-12-24).
- The Unpredictable Species: What Makes Humans Unique, Philip Lieberman 2013
- "Mind, Brain and Education, Kurt Fischer, Christina Hinton