遗传度
遗传度,又称遗传力,是育种学和遗传学使用的一种统计量,用来估计某一性状在群体中有多大比例的变异是遗传因素决定的[3],测得变异也因环境因素效应变化(含测量误差)。根据是否是受到成长家庭影响,人类的环境因素常分为“共享环境因素”和“非共享环境因素”。


遗传度通过对群体中遗传相关个体的表现型变异进行估计。遗传度是定量遗传学的重要概念,主要用于育种和行为遗传学领域(如双生子研究)。
概要
遗传度反映的是,可以归结于遗传变异的表现型变异幅度。这不是在说,个体的某一性状有多少是遗传导致的。因为即使未发生任何遗传改变,环境导致变异增加时,遗传性也可以改变。以智商为例:遗传和环境都可以影响智商,如果遗传变异增加,致使个体表现型变异上升,那么遗传度就会上升;如果环境变异下降,致使个体表现型变异下降,那么遗传度也会上升。因此,遗传度的大小关键在于遗传的相对贡献幅度。遗传度是因特定群体、特定环境而异的。
表现型受到环境的效应幅度也可能与所涉基因相关。基因对表现型的渠限化使得遗传度更加复杂,让其表达在所有环境中进行。基因型相同的个体也可能由于表型可塑性表现出不同的表现型,让有些情况下遗传度更难测量。随着近年来分子生物学的发展,使得我们可以估计环境改变对个体基因转录活动变化的影响。然而,很多基因的转录也是不受环境影响的。[4]
人们应用统计学估计遗传度的大小。由于遗传度与方差有关,它是对群体中个体变异的一种测量。遗传度可以是单变量的(单独研究一个性状),也可以是多变量的(同时研究多个性状)。这也允许对不同性状(如髮色和眼睛色彩)之间的共享遗传因素的比例进行测定。环境和遗传是相互作用的,通过遗传分析可以测试并研究这些互作。
遗传度分析的前提是:群体中存在相关变异。群体倘若不存在变异(每个个体都有或都没有),则不会存在遗传度。而实际上,所有的人类行为性状都存在变异,且几乎都存在一定的遗传度。[5]
定义
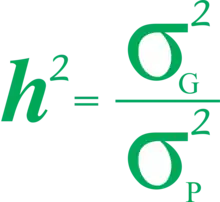
- 表现型 (P) = 基因型 (G) + 环境 (E).
性状的表现型方差(P),是下列效应之和:
- Var(P) = Var(G) + Var(E) + 2 Cov(G,E).
在设计的研究中,可以将Cov(G,E)控制为0。此时,遗传度定义为:
- .

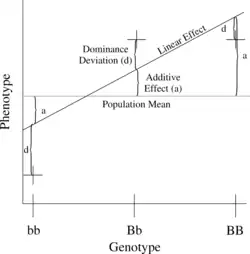
H2是广义遗传度。它反映了加性、显性、上位性和母父体效应(个体直接被父母的表型效应,如哺乳动物的产乳)对个体表现型变异的全部遗传贡献。
平均效应(加性方差)是遗传变异的主要成分,它是由于等位基因的加性作用产生的。由于父母各将基因座的一个等位基因传递给后代,亲子相似性决定于每个等位基因的平均效应。加性方差代表导致亲子相似性的遗传部分的变异。加性遗传部分占表型的方差通常称为狭义遗传度,定义为:

大写的H2指广义遗传度,小写的h2指狭义遗传度。
有的性状是二元的,比如多指或是否患病。对于这些性状,可以认为存在一个阈值,多个位点的贡献的总和此值会显现为性状,遗传度也就可以估计、选择可以建模,这称为倾向阈值模型。
加性变异对于自然选择是重要的,如果出现了可以提高动物的适应性的变异,那么对于这个性状的选择就会直接与狭义遗传度相关。如果父母的平均与产下后代父母的平均产生差别,那么群体的平均就会改变。根据观测到的选择的反应可以算出狭义遗传度。这是人工選擇或育种的原理。







估计遗传度
因为只有P可以直接观测或测量,遗传度只能从遗传和环境不同的个体的相似度来估计。对此,需要使用统计学分析方法。简单地来说,相比于亲缘关系远的个体,从遗传关系存在较大区别的个体(比如双生子、同胞、亲子)进行估计是最理想的。增加样本大小,可以使遗传度的标准误下降。
对于非人类群体,可以通过控制实验的方法收集信息。比如对于牲畜,让一只种牛与许多只母牛产下后代是很容易的。然而这种對照實驗一般无法在人类身上进行,而只能通过自然产生的关系和环境进行判定。
人类遗传度的研究通常使用收养设计,比如年幼分离,在不同环境下长大的同卵雙胞胎。他们相同的基因型可以用来分离基因和环境的效应。此设计的局限是:宫内环境仍然是共享的,而且这样的双生子数量不多。更常见的方法是,通过比较同卵和异卵双生子相似度的差异来估计遗传度。但是同卵双生子也非基因完全相同,会导致遗传度的低估。也可用双生子研究对双生子和非双生子同胞进行研究,来研究如宫内竞争的现象(比如双胎输血综合征)。
遗传度与群体中的遗传和环境因型的相对效应有关,并不是基因和环境因素对表现型贡献的绝对值。遗传度估计反映了遗传方差与环境方差的比例。
如果将遗传背景扩大,那么遗传度就会扩大(比如,可以进行远系繁殖,会提高VarG),降低环境效应(降低VarE)也可产生相似的效果。因此,相同物种的不同群体同一性状可能会显示为不同遗传度。
观察性研究或者因唤起作用(基因组通过对环境产生作用),G和E可能会共变,产生基因环境相关性。根据用于不同的估计方法,遗传因素和共享或非共享环境因素有可能会、也可能不会与遗传度产生混杂。[7]
遗传度的常见误解
遗传度通常用遗传度指数来表示,范围为0到1。0意味着研究中某一性状在该群体中完全不由遗传因素解释;1表示完全是遗传导致的。
遗传度的正确理解是,群体中由于遗传因素导致变异的比例。它不代表个体性状发育时,遗传效应的程度。比如,性格的遗传度是.6,如果说一个人60%的遗传度是从父母遗传而来,另外40%来源于环境就是不正确的。
即使是高度遗传的性状(如眼睛色彩)假定环境因素对于发育起到效应:比如适合生命生长的温度、大气层等等。“固定还是专性”是比“自然还是后天”更有助于理解区别——在特定的环境范围下,哪些性状更加“固定”(长鼻子)还是更加“专性”(对于环境改变更敏感,比如婴儿时学的语种)。
另一个有助于理解的区别是,哪些更有可能是适应(如脐带),哪些更有可能是适应的副产品(如肚脐眼),或者是由于随机变异引起的(肚脐眼的凸凹)。
估计方法
对于估计遗传度,主要有两种学派。
一种是芝加哥大学休厄尔·赖特发明的,之后被李景均(芝加哥大学)和J. L. Lush(爱荷华州立大学)推广使用。是根据对相关系数和回归的分析。休厄尔·赖特开发了路径分析的方法估计遗传度。
第二种是R. A. Fisher发明的,爱丁堡大学、爱荷华州立大学和北卡罗来纳州立大学等学校后来进一步发展了此方法。它是根据对育种研究的方差分析进行的,根据亲缘关系的组内分析进行估计。分析中使用了方差分析的多种估计方差的方法。
回归和相关的估计方法
第一种估计遗传度的学派使用回归和相关。
选择实验
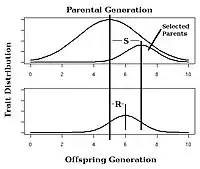
已知选择强度S(群体整体与选择的下一代父母的差别,也称为选择差数[8])和对选择的反应R(后代和上一代整体平均的差),可以借助公式h2=R/S算出相应的遗传度。
亲代间的比较
对于亲代,基本来说:

r可以被认为是关系的常数,b是回归常数,t是相关系数。
亲子回归

可以通过对父母和后代性状的比较估计遗传度。后代与父母性状回归线的斜率与性状的遗传度接近。如果只用父母一人的值,遗传度就是斜率的两倍。由于后代的值总是倾向于回归到平均,这就是迴歸分析名称的来源。DeFries Fulker分析可以用只有一人发病的双生子数据估计遗传度[9]。
同胞比较
通过对全同胞进行比较,可以简单地估计遗传度[10]。当只有加性遗传作用,全同胞的表型相关系数就是相似程度的指数:共享一半遗传因素,共享全部环境因素。全同胞表型相关性是加性遗传度的上限。半同胞方法则对同父异母、同母异父的个体进行分析。
双生子研究
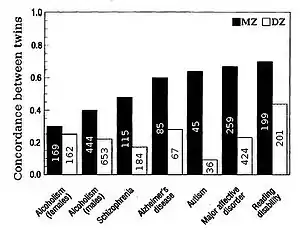
人类遗传度通常通过对同卵双生子的相似性进行比较。“双生子的研究的优点是:研究可将总体方差可以分为遗传、共享环境和独立环境,允许遗传度的精确估计”[11]。异卵双生子共享一般的基因(假定研究的性状没有选型交配),同卵双生子遗传相似性是其两倍。对遗传度的粗略估计是将同卵和异卵双生子之间相关性的差异加倍,即Falconer公式 H2=2(r(MZ)-r(DZ))。
共享环境效应c2是成长环境的共同之处。共享环境可以通过异卵双生子的相关性减去遗传度的一半进行计算(因为异卵双生子共享一半的基因),c2=DZ-1/2h2。独立环境方差e2反映了在相同环境下成长的同卵双生子的区别程度e2=1-r(MZ)。
传统双生子研究的方法的部分批评没有考虑到上面提到的方法创新和改进。
方差分析估计法
另一种估计遗传度的方法是方差分析。
基础模型
以下是Kempthorne (1957 [1969])的基础讨论。以最基础的基因模型为例,基因型为Gi的定量贡献为:

是基因型Gi的影响,是环境影响。


如果种畜和随机母畜,后代会从种畜得到一半的基因,也会从随机的母畜得到一半的基因,后代的公式为:

组内相关
如果有两组后代可以比较,第一个与同一种畜的不同后代进行比较,方差会包含遗传方差(因为母畜并非相同)和环境方差,这本认为是“误差”项。
第二组后代是对半同胞之间平均进行的比较。除了误差项,也因不同类型的半同胞增添了一项,组间相关性为:

因为环境效应是相互独立的。
方差分析
对于包括个种畜和其平均的个后代的实验,我们可以使用下面的方法计算方差:


| 来源 | 自由度 | 平均的平方 | 期望平均平方 |
|---|---|---|---|
| 种畜群体之间 | |||
| 种畜群体之内 |






一项是半同胞之间的组内相关。我们可以容易地算出。可以从个体之间预期的平均平方的关系计算出预期平均平方。

















对选择的反应
在对植物和动物的人工選擇中,已知狭义遗传度 可以用育种公式估计对选择的反应[15]:


公式中,对选择的反应(R)定义为亲代到子代的平均差异,选择差异(S)定义为亲代平均与部分亲代的平均差异。
比如,想象一个植物育种师想要提高每穗玉米平均粒数。亲代平均产下100粒,而部分亲代产下120粒。如果h2等于0.5,那么下一代每穗会平均多产下0.5(120-100) = 10粒,平均产下共110粒。
请注意,只有当基因型和环境干扰服从正态分布,比例才与相等。

争议
遗传度突出的批评者包括Steven Rose[16]、Jay Joseph[17]和Richard Bentall,注意力集中于行为科学和社会科学对遗传度估计的解读。Bentall称,为算出高分,遗传度分数通常通过违背常理的方式计算,这使得人们转移对研究发现的其他因素的注意力,比如儿童虐待导致成年的精神错乱[18][19]。
对测量遗传度的争议一般是对双生子研究的。分子遺傳學研究提供的遗传度下限一般会低于分子遺傳學研究的现象称为丢失的遗传度问题[20]。Eric Turkheimer认为新近的分子学研究支持了传统对双生子研究的解读[20],然而基因与行为之间的关系的解释仍然不清楚[21]。Turkheimer主张,基因与环境都是可以遗传的,不同的环境下遗传贡献不同,注意于遗传度会分散对其他重要因素的关注[22]。然而,一般来说,遗传度的概念还是受到广泛应用的[14]。
参考资料
参考
- 中山大学中文系. 《古汉语基础知识》编写组. . 广东人民出版社. 1979: 107 [2020-09-12]. (原始内容存档于2021-07-01) (中文).
- 李盟编,DNA密码,中国言实出版社,2012.04,第133页
- Wray, Naomi; Visscher, Peter. . Nature Education. 2008, 1 (1): 29 [2015-07-24]. (原始内容存档于2015-08-02).
- Wills, C. . Journal of Heredity (Book Review). 2007, 98 (4): 382–382. doi:10.1093/jhered/esm035.
- review of: Hartl, Daniel L.; Clark, Andrew G. Sunderland, MA: Sinauer and Associates. 2007: xv + 652. ISBN 0-87893-308-5. 缺少或
|title=为空 (帮助)
- review of: Hartl, Daniel L.; Clark, Andrew G. Sunderland, MA: Sinauer and Associates. 2007: xv + 652. ISBN 0-87893-308-5. 缺少或
- Turkheimer, Eric. (PDF). Current Directions in Psychological Science. October 2000, 9 (5): 160–164 [2013-10-29]. ISSN 0963-7214. doi:10.1111/1467-8721.00084. (原始内容存档 (PDF)于2013-10-19).
- Kempthorne 1957
- Cattell RB. . Psychol Rev. 1960, 67 (6): 353–372. PMID 13691636. doi:10.1037/h0043487.
- Kempthorne 1957,第507頁; or Falconer & Mackay 1995,第191頁, for example.
- Defries, J. C.; Fulker, D. W. . Behavior Genetics. September 1985, 15 (5): 467–473. PMID 4074272. doi:10.1007/BF01066239.
- Falconer, Douglas S.; Mackay, Trudy F. C. 4th. 朗文出版社. December 1995. ISBN 978-0582243026.
- Gielen, M., Lindsey, P.J., Derom, C., Smeets, H.J.M., Souren, N.Y., Paulussen, A.D.C., Derom, R., & Nijhuis, J.G. (2008) "Modeling Genetic and Environmental Factors to IncreaseHeritability and Ease the Identification of Candidate Genes for Birth Weight: A Twin Study". Behavioral Genetics. 38(44-54):45. DOI 10.1007/s10519-007-9170-3
- Luciano, M.; Batty, G. D.; McGilchrist, M.; Linksted, P.; Fitzpatrick, B.; Jackson, C.; Pattie, A.; Dominiczak, A. F.; Morris, A. D.; Smith, B. H. . Intelligence. May–June 2010, 38 (3): 304–313. doi:10.1016/j.intell.2010.03.002.
- Hill, W. G.; Goddard, M. E.; Visscher, P. M. MacKay, Trudy F. C. , 编. . PLOS Genetics. 2008, 4 (2): e1000008. PMC 2265475
 . PMID 18454194. doi:10.1371/journal.pgen.1000008.
. PMID 18454194. doi:10.1371/journal.pgen.1000008. 
- Visscher, P. M.; Hill, W. G.; Wray, N. R. (PDF). Nature Reviews Genetics. April 2008, 9 (4): 255–266 [2017-07-27]. PMID 18319743. doi:10.1038/nrg2322. (原始内容存档 (PDF)于2016-03-24).
- Plomin, R., DeFries, J. C., & McClearn, G. E. (1990). Behavioral genetics. New York: Freeman.
- Rose, Steven P R. . International Journal of Epidemiology. 2006, 35 (3): 525–527. PMID 16645027. doi:10.1093/ije/dyl064.
- Jay Joseph, The Gene Illusion (New York: 阿尔戈拉, 2004), esp ch 5 (页面存档备份,存于).
- Richard P Bentall, Doctoring the Mind: Is Our Current Treatment of Mental Illness Really Any Good? (New York: New York University Press, 2009), p 123–127 (页面存档备份,存于).
- Melanie McGrath, "Doctoring the Mind: Review" (页面存档备份,存于), The Telegraph, 2009 Jul 5.
- Turkheimer, Eric. . Research in Human Development. 2011, 8 (3–4): 227–241. doi:10.1080/15427609.2011.625321.
- Eric Turkheimer, "Genetic prediction", Hastings Center Report, 2015 Sep/Oct;45(S1):S32–S38.
- Jay Joseph, The Trouble with Twin Studies: A Reassessment of Twin Research in the Social and Behavioral Sciences (页面存档备份,存于) (New York & Hove: 羅德里奇, 2015), esp p 81 chapter summaries (页面存档备份,存于).
图书
- Falconer, D. S. and T. Mackay. 1996. Introduction to Quantitative Genetics, Ed. 4. Longman, Essex, England.
- Kempthorne, O (1957 [1969]) An Introduction to Genetic Statistics. John Wiley. Reprinted, 1969 by Iowa State University Press.
延伸阅读
- Lynch, M. & Walsh, B. 1996. Genetics and Analysis of Quantitative Traits. Sinauer Associates. ISBN 0-87893-481-2.
- Johnson, Wendy; Penke, Lars; Spinath, Frank M. . European Journal of Personality. 2011, 25 (4): 287–294 [2013-12-15]. ISSN 0890-2070. doi:10.1002/per.835. (原始内容存档于2017-08-12).