雙腔龍屬
雙腔龍屬(學名:Amphicoelias)是蜥腳下目梁龍科恐龍的一個屬。雙腔龍的學名是由古希臘文的「amphi」(意即「兩面」)及「koilos」(意即「空腔」或「空洞」)而來,所以中文翻譯為雙腔龍。根據唯一的骨頭化石,長度估計約達40-60公尺,而體重可能約為122公噸,可比現今最重的動物藍鯨。但在1870年代描述這種恐龍後不久,這塊唯一的骨頭就遺失了,存有的證據就只有一些插圖及紀錄。雙腔龍的化石,都發現於莫里遜組的第6地層帶[1]。
| 双腔龙属 化石时期:侏罗纪晚期, | |
|---|---|
 | |
| 高雙腔龍的正模標本 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 亚目: | †蜥脚形亚目 Sauropodomorpha |
| 下目: | †蜥腳下目 Sauropoda |
| 科: | †梁龙科 Diplodocidae |
| 属: | †双腔龙属 Amphicoelias Cope, 1878 |
| 模式種 | |
| 高雙腔龍 Amphicoelias altus Cope, 1878 | |
2018年肯尼斯·卡本特(Kenneth Carpenter)將易碎雙腔龍(A. fragillimus)命名為新屬極巨龍屬,並將其分類為雷巴齊斯龍科。
描述
雙腔龍屬的模式種是高雙腔龍(A. altus),是由愛德華·德林克·科普(Edward Drinker Cope)於1877年12月根據一個不完整的骨骼所命名,化石包含了2節脊椎、一個恥骨、及一個股骨,但要到1878年才公佈[2]。科普也命名第二個種,寬雙腔龍(A. latus)。目前古生物學界認為寬雙腔龍是高雙腔龍的異名。在1921年,亨利·費爾費爾德·奧斯本(Henry Fairfield Osborn)與C.C. Mook將一塊肩胛骨、一塊鳥喙骨、尺骨及一顆牙齒編入高雙腔龍。這些化石顯示了雙腔龍與梁龍的高度相似性,及少數的主要差異,如雙腔龍的前肢在比例上較梁龍為長[3]。雙腔龍的股骨也是異常的長、修長,及在橫切面呈圓形,圓形橫切面曾一度是雙腔龍的獨有特徵,後來在一些梁龍標本中也有出現[4]。高雙腔龍在體型上與梁龍相近,估計約有25公尺長[5]。大部分研究人員根據化石特徵的差異,認為雙腔龍、梁龍是不同屬;少數研究人員則提出雙腔龍可能是梁龍的次異名[6]。

雙腔龍的第三個種易碎雙腔龍(A. fragillimus),只是一個高1.5公尺的不完整神經弓,位於背部脊椎的倒數第一或倒數第二節(D10或D9),在生前這骨頭應有2.7公尺長。除了這部分脊椎,科普的紀錄中也有一塊「股骨的末端」,位於該巨大脊椎數十米之外,而這個沒有被描述的股骨也被認為是來自於同一個體[4]。
在2010年的一份專題論文,提出雙腔龍屬的第四個種。Henry Galiano、Raimund Albersdorfer將發現於懷俄明州大角盆地、私人收藏的數個完整標本,非正式命名為「"A. brontodiplodocus"」。他們認為莫里遜組的大部分梁龍科化石,其實是雙腔龍的不同生長階段、或是不同性別,因此種名綜合雷龍與梁龍[7]。但許多人質疑不同生長階段、不同性別雙腔龍的理論[8]。原始研究人員也不再主張這個理論。
歷史
易碎雙腔龍是由古生物學家愛德華·德林克·科普(Edward Drinker Cope)所聘請的化石收藏家Oramel Lucas在1877年所挖掘的。Lucas在美國科羅拉多州的卡農城北方的花園公園發現這個部分脊椎(神經弓及神經棘),接近發現圓頂龍的礦場。這個部分脊椎的保存狀況很差,但卻異常的大,高度有1.5公尺高,估計完整的脊椎可達2.7公尺高[4]。他在1878年把這個標本運送給科普,而科普以此作為新種易碎雙腔龍的正模標本(編號AMNH 5777),並於同年8月公佈[9]。牠的種名是由古希臘文的「fragillimus」而來,意為「非常易碎」,意指神經弓的椎板非常薄。從科普發掘時的紀錄得知,這個標本從發現圓頂龍的礦場南面的山上被發現。科普指該位置是屬於達科他組(Dakota Formation),年代為白堊紀中期,而在同一地層的圓頂龍則是屬於莫里遜組(Morrison Formation),年代可追溯至晚侏羅紀的提通階[10]。一塊巨大的股骨化石也在附近被發現,被推論可能也是屬於易碎雙腔龍。
遺失的巨型化石
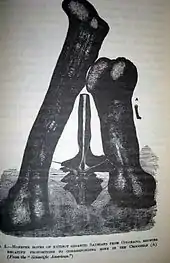
當討論最大的恐龍時,一般都會忽略易碎雙腔龍的巨大骨頭,這是由於不同的報告都指其脊椎及股骨的下落不明,而嘗試尋找的計劃都失敗[3][9]。於2006年,肯尼思·卡彭特(Kenneth Carpenter)指出易碎雙腔龍標本失蹤的可能原因:由於科普在其描述中指該神經弓非常易碎,而當時並沒有使骨頭硬化及保存化石的相關技術;科普的對手,奧塞內爾·查利斯·馬什(Othniel Charles Marsh)在1880年代首次採用此一技術。而保有易碎雙腔龍化石的石礦是嚴重侵蝕的泥岩,使得該化石容易被粉碎及形成小型、不規則的碎片。所以,它可能在科普描繪完後就被粉碎了,所以科普的插圖就只有一個視圖,而不是一般的多個視圖[4]。

於1994年,有科學家嘗試去找出易碎雙腔龍的原有石礦,當中使用了雷達來探測地下藏有的骨頭影像。這個嘗試最後失敗,是由於化石化的泥岩與週邊的岩石同一密度,以致不能分辨出兩者。另一個地形研究指出藏有化石的地層經已受嚴重侵蝕,可能在發掘易碎雙腔龍的脊椎及股骨後,該地的大部分的骨骼已經侵蝕、消失[4]。
由於易碎雙腔龍化石的特殊尺寸及神秘失蹤,科普的原始描述就引來了懷疑,有些人更指出科普可能是在量度上出現了排版印刷之誤。但是卡彭特指出科普的文章充滿根據,也顯示他是一位出色的古生物學家。而發現化石的時間正好是化石戰爭,當時科普的競敵奧塞內爾·查利斯·馬什(O.C. Marsh)卻從沒有對易碎雙腔龍提出懷疑。而馬什亦曾僱用偵探監視科普的發現,可能對易碎雙腔龍骨頭的巨大有了確定[4]。而奧斯本、C.C. Mook、 John S. McIntosh等不同的學者亦都接受科普的資料[3][9]。
體型
(易碎雙腔龍現已為無效種,故該種的體型估計應納入易碎極巨龍之研究)


若要估計易碎雙腔龍(現為易碎極巨龍)的大小,就須假設牠與梁龍科的相對比例相似,以較多瞭解的梁龍科的骨頭作縮放比例。在科普原先的紀錄,他以此來估計易碎雙腔龍(現為易碎極巨龍)的股骨大小。科普發現在其他的蜥腳下目恐龍,尤其是高雙腔龍及至高圓頂龍,牠們的股骨都是背部最高脊椎的兩倍高度,因而估計易碎雙腔龍(現為易碎極巨龍)的股骨為3.7公尺高[11]。於1994年,葛瑞格利·保羅(Gregory S. Paul)利用相關的梁龍作為參考,估計易碎雙腔龍(現為易碎極巨龍)的股骨大小為3.1到4米長[5]。於2006年,卡彭特就同樣利用梁龍重新評估易碎雙腔龍(現為易碎極巨龍)的股骨為4.3到4.6公尺[4]。卡彭特嘗試就整頭易碎雙腔龍(現為易碎極巨龍)的體型作出評估,但發現梁龍科不同種的體型差異,會影響其估計結果。假設易碎雙腔龍(現為易碎極巨龍)與梁龍有著同一的比例,牠的體型就是58公尺,與保羅在1994年的估計範圍(40到60公尺)吻合。卡彭特也指出易碎雙腔龍(現為易碎極巨龍)的最小估計值,亦比其他蜥腳下目更為巨大,如超龍(32.5公尺)、腕龍科的波塞東龍(34公尺)及泰坦巨龍類的阿根廷龍(30公尺)。卡彭特版本的易碎雙腔龍身體比例中,頸部長16.75公尺,體長9.25公尺,尾巴長32公尺。整個前肢高達5.75公尺,而後肢高7.5公尺,背部最高點的高度為9.25公尺[4]。而目前平均最長的現存生物藍鯨,則長約30公尺[12]。
分類
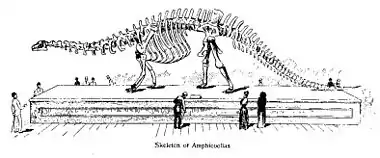
在1878年,科普將他的發現公佈於《美國博物學家》(The American Naturalist)的兩份期刊,並新命名為雙腔龍屬。他把雙腔龍屬編入獨自的雙腔龍科(Amphicoeliidae);由於超過50年沒有被科學研究使用,雙腔龍科目前是個遺失名。雖然只有部分的化石碎片,雙腔龍常被歸類於梁龍科。近年有些學者則將雙腔龍分類在梁龍超科下,或是梁龍科的「分類未定屬」[13]。雙腔龍屬的第一個被命名種是高雙腔龍(正模標本AMHD 5764),是由科普在1877年發現。雖然只有部分的身體骨骼,但有足夠特徵可以建立此屬的一些特徵。高雙腔龍的化石較為完好,但就比易碎雙腔龍為小。在1878年,科普命名第三個種,寬雙腔龍(Amphicoelias latus)[2]。
在1921年,奧斯本與C.C. Mook指出易碎雙腔龍其實只是非常大的高雙腔龍,是高雙腔龍的異名,並提出寬雙腔龍是高雙腔龍的相同種;在1998年,John S. McIntosh也同意這個看法[3][9]。但是卡彭特指出,牠們在脊椎的構造上有所差異,足夠顯示牠們是不同的種,或甚至不同的屬。另外,卡彭特也提出只根據原有標本,很難去確定易碎雙腔龍的有效性。
在2007年,John Foster提出高雙腔龍、梁龍屬的特徵差異並不明顯,兩者細微差異可能是相同物種的個體差異。John Foster因此提出雙腔龍可能是梁龍的次異名。如果屬實,較早命名的雙腔龍將擁有優先權,梁龍將失去有效名;如同較早命名的迷惑龍、較著名但無效的雷龍(雷龙现已正名为有效种,是与迷惑龙相似但不同的另一种恐龙)[6]。
古生物學

為何包括雙腔龍在內的蜥腳下目恐龍會有著這麼巨大的體型?卡彭特指出這種體型是在蜥腳類恐龍的早期演化階段出現的,而在晚三疊紀時期就已存在著巨大的物種,顯示蜥腳類恐龍的祖先有著巨大化的演化壓力。巨大的草食性哺乳動物(如象及犀牛),都顯示巨大的草食性動物會有較高的消化效率。由於大型的動物有較長的消化系統,食物會留在身體較長時間,容許牠們在低營養價值的食物來源下生存。在小腸內亦會有大量的發酵室,容許微生物積聚及發酵植物,協助消化。在其演化的歷史中,蜥腳下目主要都是在半乾燥及乾旱的環境發現,而在旱季有著季節性的食物營養價值下降。雙腔龍的環境是大草原,而現今巨大草食性動物生存於乾燥環境,支持了以上的說法。其他巨大體型的優勢則被認為是次要的,如對獵食動物的相對抵抗力、低能量消耗、或長壽等[4]。
雙腔龍生活的莫里遜組環境類似現今的大草原,由於草要到上白堊紀才出現,蕨類可能是莫里遜組的主要植物及雙腔龍的食物來源。雖然有研究指出低卡路里的蕨類不適合作為食物[14],但是蜥腳下目恐龍的消化系統是適合大量處理這些低營養價值的食物[4]。而這些恐龍的巨大體型,顯示應有達20-30米高的巨大樹木食物來源,與大草原環境有所矛盾。但是,由於巨大的樹木需要更多的水份,它們可能只有很少數量,及沿河流及小峽谷生長。而更有研究指出雙腔龍是以這些樹木作為遮蔭,日間可以令身體涼快,而夜間才在草原攝食[4]。
參考資料
- (英文)Foster, J. (2007). "Appendix." Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. pp. 327–329.
- (英文)Cope, E.D. (1878a). "On the Vertebrata of the Dakota Epoch of Colorado." Proceedings of the American Philosophical Society, 17: 233-247.
- (英文)Osborn, H.F., and Mook, C. C. (1921). "Camarasaurus, Amphicoelias and other sauropods of Cope." Memoirs of the American Museum of Natural History NS, 3(3): 249-387.
- (英文)Carpenter, K. (2006). "Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus." In Foster, J.R. and Lucas, S.G., eds., 2006, Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36: 131-138. (页面存档备份,存于)
- (英文)Paul, G.S.(1994a). "Big sauropods - really, really big sauropods." The Dinosaur Report, The Dinosaur Society, Fall, p. 12-13.
- (英文)Foster, J. (2007). Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press.
- (英文)Galiano, H. and Albersdorfer, R. "A new basal diplodocid species, Amphicoelias brontodiplodocus, from the Morrison Formation, Big Horn Basin, Wyoming, with taxonomic reevaluation of Diplodocus, Apatosaurus, and other genera. (页面存档备份,存于)" Dinosauria International, LLC. 44pp. Published online 2010.
- (英文)Mike Taylor. . Sauropod Vertebra Picture of the Week. [2012-09-16]. (原始内容存档于2011-12-27).
- (英文)McIntosh, J.S. (1998) "New information about the Cope collection of sauropods from Garden Park, Colorado." In Carpenter, K., Chure, D. and Kirkland, J.I., eds., The Morrison Formation: an interdisciplinary study: Modern Geology, 23: 481-506.
- (英文)Turner, C.E., and Peterson, F. (1999) "Biostratigraphy of dinosaurs in the Upper Jurassic Morrison Formation of the Western Interior, U.S.A." In. Gillette, D., ed., Vertebrate Paleontology in Utah: Utah Geological Survey Miscellaneous Publication, 99(1): 77-114.
- (英文)Cope, Edward Drinker. . American Naturalist. 1878b, 12 (8): 563–564 [2010-10-08]. doi:10.1086/272176. (原始内容存档于2009-10-25).
|author=和|last=只需其一 (帮助) - (英文)J. Calambokidis and G. Steiger. . Voyageur Press. 1998. ISBN 0-89658-338-4.
- (英文)Wilson, J.A., and Smith, M. (1996). "New remains of Amphicoelias Cope (Dinosauria: Sauropoda) from the Upper Jurassic of Montana and diplodocoid phylogeny." Journal of Vertebrate Paleontology, 16(3 Suppl.): 73A.
- (英文)Engelmann, G.F., Chure, D.J., and Fiorillo, A.R. (2004). "The implications of a dry climate for the paleoecology of the fauna of the Upper Jurassic Morrison Formation." In Turner, C.E., Peterson, F., and Dunagan, S.P., eds., Reconstruction of the extinct ecosystem of the Upper Jurassic Morrison Formation: Sedimentary Geology, 167: 297-308