埃德蒙頓龍屬
埃德蒙頓龍屬或愛德蒙托龍(學名:Edmontosaurus),是鴨嘴龍科下的一屬恐龍,可達13米長,平均體重約6-9噸,最大体重11.6吨,是最大的鴨嘴龍科之一,也是最晚進化出來的鸭嘴龙類之一。愛德蒙托龍生活於上白堊紀的坎潘階晚期至馬斯垂克階晚期的美國地獄溪組及加拿大,距今約7300萬到6600萬年前,以加拿大發現的化石數量為最多,牠的名稱也是以加拿大的艾伯塔省來命名的,這些愛德蒙托龍和暴龍、甲龍、腫頭龍以及三角龍生活在同一個地質年代。
| 埃德蒙顿龙属 | |
|---|---|
 | |
| 牛津大學自然史博物館中展覽的埃德蒙頓龍骨骼 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | †鸟臀目 Ornithischia |
| 亚目: | †鸟脚亚目 Ornithopoda |
| 科: | †鴨嘴龍科 Hadrosauridae |
| 亚科: | †栉龙亚科 Saurolophinae |
| 族: | †埃德蒙顿龙族 Edmontosaurini |
| 属: | †埃德蒙顿龙属 Edmontosaurus Lambe, 1917 |
| 模式種 | |
| †帝王埃德蒙頓龍 Edmontosaurus regalis Lambe, 1917 | |
| 其他種 | |
| 異名 | |
埃德蒙頓龍的第一個化石發現於艾伯塔省的馬蹄峽谷組。模式種是帝王埃德蒙頓龍(E. regalis),是由勞倫斯·賴博(Lawrence Lambe)在1917年命名。目前還有其他已命名種,其中最著名的種是E. annectens,最初是由奧塞內爾·查利斯·馬什(Othniel Charles Marsh)在1892年命名,當時為破碎龍的一種(Claosaurus annectens),後曾被長期歸類於鴨龍,直到近年才被歸類於埃德蒙頓龍。
特徵

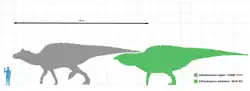
埃德蒙頓龍目前已發現數個標本[1][2][3][4]。如同其他鴨嘴龍類,埃德蒙頓龍的頭部前段平坦、寬廣,口鼻部類似鴨子,雄性具小型頭冠,尾巴長而窄。前肢短於後肢短,但前肢亦有足夠長度,仍適宜行走。
完全成長的埃德蒙頓龍可達9公尺長,一些較大的標本則達12公尺[5]到13公尺長[6]。體重約4.0噸,是最重的鴨嘴龍科之一[7]。帝王埃德蒙頓龍是體型最大的種;在2004年,傑克·霍納(Jack Horner)等人提出科氏大鴨龍是E. annectens的異名,但很少其他研究引用這個分類法[7]。模式標本(編號NMC 2288)的身長估計為9到12公尺[8]。E. annectens目前已發現兩個標本,身長分別為8公尺、8.92公尺[8][9]。但是,有一個可能屬於E. annectens的標本,身長被估計為約12公尺[10]。薩斯喀徹溫埃德蒙頓龍的身長則為7到7.3公尺[4]。
頭骨

埃德蒙頓龍的頭骨長度約1公尺,帝王埃德蒙頓龍的頭骨較長,而E. annectens的頭骨較短[8]。埃德蒙頓龍的頭部側面略呈三角形[1],而且沒有骨質頭冠[11]。若從頭骨的上方看,頭部的前端與後段較寬,中段狹窄。埃德蒙頓龍具有缺乏牙齒的喙狀嘴,由角質組織覆蓋[7]。根據位在森肯貝格博物館的「糙齒龍木乃伊」標本,埃德蒙頓龍的喙狀嘴角質部分至少有8公分長[12]。埃德蒙頓龍的鼻孔大,鼻孔周圍的骨頭凹陷[13]。根據「糙齒龍木乃伊」標本,埃德蒙頓龍的眼眶具有鞏膜環(Sclerotic rings)[14]。
埃德蒙頓龍只有上頜骨與齒骨具有牙齒。新的牙齒會不斷的生長來取代脫落的牙齒,一顆牙齒需要約一年的時間來生長[15]。牙齒排列成形成數十列齒系(Dental batteries),每列齒系有至少6顆牙齒;而齒系的數量,依該物種而變動[4]。帝王埃德蒙頓龍的每塊上頜骨有51到53列齒系,每塊齒骨有48到49列齒系,而上頜的牙齒較下頜牙齒狹窄。E. annectens的每塊上頜骨有43列齒系,每塊齒骨有36列齒系。薩斯喀徹溫埃德蒙頓龍的每塊上頜骨有52列齒系,每塊齒骨有44列齒系[11]。
身體
埃德蒙頓龍的脊椎數量,因不同種而有不同變化。帝王埃德蒙頓龍有13節頸椎、18節背椎、9節薦椎、尾椎的數量則不清楚[11]。一個E. annectens標本被認為具有19節背椎、85節尾椎[11]。由於其他鴨嘴龍類的尾椎多為50到70節之間[7],上述估計值可能過度誇大。埃德蒙頓龍的背部與尾巴脊椎骨具有骨化肌腱,可使背部與尾巴保持僵硬[16][17]。這些骨化肌腱被認為提供四肢以外的額外支撐脊椎[16]。肩胛骨長而平坦,類似刀狀,與脊椎平行。骨盆包含三個骨頭:長的腸骨、長而薄的坐骨、板狀的恥骨。骨盆的結構允許埃德蒙頓龍以二足方式站立。而9節癒合的薦椎也使骨盆牢固[4]。
與後肢相比,埃德蒙頓龍的前肢較短、較細。肱骨具有大型的三角嵴(Deltopectoral crest),可供胸鎖三角肌肉附著,尺骨與橈骨纖細。上臂與前臂的長度相當。手腕只有兩塊小型腕骨。手掌具有四指,缺乏俗稱的拇指。前三指的長度相當,生前可能由肉質組織連接。前兩指具有蹄狀指爪,外表由皮膚覆蓋。最後一指的長度較短,可做出獨立於其他三指的動作。股骨筆直而粗壯,後側具有明顯的第四粗隆部[4]。連接大腿與臀部、尾巴的肌肉,會連接至第四粗隆部;當動物前進時,可使尾巴發生平衡的功能[14]。腳掌具有三趾,缺乏第一與第五趾。腳趾具有蹄狀趾爪[4]。
分類
|
埃德蒙頓龍屬於鴨嘴龍科,一群具有鴨嘴型態口鼻部的植食性恐龍,生存於白堊紀晚期。埃德蒙頓龍屬於其中的鴨嘴龍亞科,其頭部缺乏中空頭冠。鴨嘴龍亞科還包含:格里芬龍、慈母龍、短冠龍、納秀畢吐龍、櫛龍、冠長鼻龍、以及原櫛龍[7]。埃德蒙頓龍是大鴨龍的近親[19];但也有科學家提出大鴨龍其實是頭骨在化石化過程中遭到擠壓變形的後果,埃德蒙頓龍與大鴨龍是相同物種[7]。
埃德蒙頓龍屬於鴨嘴龍亞科,但其演化位置仍未確定。早期的系統發生學研究大多將埃德蒙頓龍與鴨龍,共同歸類於鴨嘴龍亞科的眾多演化支之一,例如理查·史旺·魯爾( Richard Swann Lull)與尼爾達·萊特(Nelda Wright)在1942年提出的北美洲鴨嘴龍科專題論文[20]。在90年代,科學家採用親緣分支分類法研究埃德蒙頓龍的演化關係,發現埃德蒙頓龍與中國的山東龍共同屬於埃德蒙頓龍類;鴨嘴龍亞科還包含以下次分類:尖頭冠的櫛龍類、鼻部拱起的格里芬龍類、以及短冠龍類[19]。在2007年,特里·蓋茨(Terry Gates)與史考特·山普森(Scott Sampson)提出類似的研究,埃德蒙頓龍是櫛龍與原櫛龍的近親,是格里芬龍、短冠龍、慈母龍的遠親[18]。在2004年,傑克·霍納(Jack Horner)等人提出不同結果的研究,他們認為埃德蒙頓龍是格里芬龍、短冠龍類的近親,而離櫛龍的關係較遠[7]。由於目前缺乏鴨嘴龍亞科的演化關係研究,這兩種不同版本的研究仍然未解。
發現與種
埃德蒙頓龍是由勞倫斯·賴博(Lawrence Lambe)於1917年所命名,其化石是於艾伯塔省馬蹄峽谷組中發現,模式種是帝王埃德蒙頓龍(E. regalis)。在1892年,奧塞內爾·查利斯·馬什(Othniel Charles Marsh)建立破碎龙的Claosaurus annectens,但後來一度分類於鴨龍(A. annectens)、目前屬於埃德蒙頓龍(E. annectens)。另外,查爾斯·斯騰伯格(Charles Mortram Sternberg)於1926年命名了萨斯喀彻温強龍(Thespesius saskatchewanensis),後來也一度分類於鴨龍(A. saskatchewanensis),但現在亦被認為是屬於埃德蒙頓龍,更名為萨斯喀彻温埃德蒙顿龙(E. saskatchewanensis)。
鴨嘴龍科中的鴨龍(Anatosaurus)意為「鴨蜥蜴」。鴨龍曾有五個種,模式種是Anatosaurus annectens,種名意為「連接」。模式種被歸類於埃德蒙頓龍後,鴨龍這個名稱被棄置,而成為埃德蒙頓龍的次異名。而薩斯喀徹溫強龍先是被歸類於薩斯喀徹溫鴨龍(A. saskatchewanensis),後被編入埃德蒙頓龍中,成為了萨斯喀彻温埃德蒙顿龙。埃德蒙頓鴨龍(A. edmontonensis或A. edmontoni)亦被認為與E. annectens是相同物種。兩個另外的鴨龍種,長頭鴨龍(A. longiceps)及科氏鴨龍(A. copei)被發現與埃德蒙頓龍不同,而被建立為新屬大鴨龍。
古生物學
生活環境

埃德蒙頓龍與暴龍生存於相同時期的相同環境,其中一個在丹佛自然科技博物館展覽的成年標本,顯示牠的尾巴曾被獸腳類恐龍所咬。尾部的第13到17節脊椎的神經棘具有明顯的牙齒痕跡,其中一個神經棘斷裂,而其他神經棘彎曲,顯示曾經被肉食性動物從後方攻擊過。由於這個部位的高度至少有2.9公尺,攻擊者應是種體型巨大的動物,而從該化石的發現地區來判斷,該地區的唯一大型肉食性動物是暴龍。骨骼顯示了這個傷口正在痊癒,並有感染的跡象,顯示這頭埃德蒙頓龍在事後應該仍然生存。研究這個標本的肯尼思·卡彭特(Kenneth Carpenter),發現左臀也有個痊癒的骨折跡象,由於已經完全康復,發生時間應較尾巴的傷口早。卡彭特研判這隻埃德蒙頓龍因為左臀的傷口而行動不便,因此成為暴龍的攻擊目標,造成尾巴的新傷口。卡彭特更提出,這隻埃德蒙頓龍成功逃過暴龍的攻擊,或是曾用尾巴抵抗暴龍的嘴部攻擊[21]。另一個發現於南達科塔州的E. annectens標本,身長約7.6公尺,下頜則具有小型獸腳類恐龍的齒痕。其中部分齒痕已經初步痊癒。Michael Triebold認為曾有小型獸腳類恐龍攻擊埃德蒙頓龍的喉嚨,雖然這隻埃德蒙頓龍在攻擊中存活下來,但不久後因為這個傷口而死亡[22]。
在馬蹄峽谷組中,埃德蒙頓龍生存於接近岸邊的環境,而亞冠龍、櫛龍則生存於較內陸的環境[23]。通常不會在一個地點同時發現埃德蒙頓龍與櫛龍的化石[24]。馬蹄峽谷組的帝王埃德蒙頓龍化石,集中在最下層的5個地點,最上二層沒有埃德蒙頓龍的化石[25]。科學家原本估計馬蹄峽谷組的年代約為馬斯垂克階早期[26];但馬蹄峽谷組的範圍很廣,最新的估計是坎潘階晚期,約7,300萬年前[27]。
社會行為
經常在許多屍骨層發現埃德蒙頓龍等鴨嘴龍類的化石,顯示牠們是群居動物[7]。在2007年,科學家歸納出四個具有埃德蒙頓龍化石的屍骨層,分佈於阿拉斯加州的王子溪層、亞伯達省的馬蹄峽谷組、南達科塔州的地獄溪組、以及懷俄明州的蘭斯組[28]。
其中,懷俄明州蘭斯組的屍骨層,發現於石灰岩與泥岩中,面積達1平方公里,而埃德蒙頓龍的化石集中於40公頃的地點內。科學家估計該地有10,000到25,000隻埃德蒙頓龍的化石。科學家也估計這些埃德蒙頓龍聚集於淡水的岸邊,然後死後的屍體被沖積到較深的水域[29]。
在亞伯達省的屍骨層,經常發現艾伯塔龍與蜥鳥盜龍的齒痕[30]。在另一個亞伯達省的化石地點,則在埃德蒙頓龍與櫛龍的化石發現懼龍的齒痕[24]。
埃德蒙頓龍的頭部缺乏骨質頭冠,而鼻孔周圍的骨頭凹陷,科學家推測其鼻孔在生前覆蓋者膨脹的氣囊,可能具有視覺辨識、或發聲的功能[13]。埃德蒙頓龍的體型可分為粗壯、瘦長兩種類型,可能是兩性異形的特徵,但科學家仍無法確定[31]。
由於埃德蒙頓龍的分佈廣泛,北至阿拉斯加州,南到科羅拉多州,其中包含北極區,因此有科學家提出埃德蒙頓龍可能具有遷徙過冬的習性。在2008年,Phil R. Bell、Eric Snively提出一份關於恐龍遷徙的研究,認為大部分生存於極區的恐龍可能有過冬的習性,包含:獸腳亞目、蜥腳下目、甲龍下目、稜齒龍類、鴨嘴龍科;而帝王埃德蒙頓龍可能會遷徙達2,600公里的路程,每小時大約移動2到10公里[32][33]。
木乃伊化石


目前已有數個埃德蒙頓龍標本被發現化石化的皮膚痕跡,其中以1908年發現的「糙齒龍木乃伊」(Trachodon mummy)[34][35]與在1999年發現的恐龙木乃伊「達科塔」標本最為有名,但後者還沒有正式的發表研究[36][37]。由於上述皮膚標本,可知埃德蒙頓龍覆蓋者類似爬行動物的鱗片。
在1908年,化石收集者查爾斯·斯騰伯格(Charles Hazelius Sternberg)與他的三個兒子,在懷俄明州康弗斯縣發現了一個保存良好的埃德蒙頓龍標本(編號AMNH 5060),屬於E. annectens,同時保存了化石化的皮膚痕跡,該標本被稱為「糙齒龍木乃伊」(Trachodon mummy)。雖然斯騰伯格替英國自然科學博物館工作,但他將這個標本以2,000美金賣給美國自然歷史博物館,由亨利·費爾費爾德·奧斯本(Henry Fairfield Osborn)在1912年研究。
這個標本被幾乎完整的保存下來,上附有一些肌肉。皮膚痕跡覆蓋者口鼻部、大部份頸部與胸部、部份四肢[12]。尾巴與部分後肢因為侵蝕作用而消失[35]。但是在處理化石的過程中,部分頸部與手掌隨者周圍石塊而移除了[34]。奧斯本推測該隻埃德蒙頓龍死於乾涸的河床,被洪水沖刷到含豐富黏土的沉積層,因此皮膚得以保存下來[35]。
這個標本的表皮薄,鱗片由小型、不互相覆蓋的鱗片構成[34],類似希拉毒蜥[12]。鱗片可分為兩種型態:小型鱗片的直徑為1到3公厘,表面凸,沒有特定的排列方式;大型鱗片的直徑不大於5公厘(前肢部份的大型鱗片直徑可達1公分),表面平坦,形狀呈多角形,大型鱗片由小型鱗片包圍者。
一個發現於北達科塔州的E. annectens幼年標本,尾巴部份保存了皮膚痕跡,皮膚分佈者硬塊與結節、橢圓形的重疊鱗片、有溝紋的鱗片,以及長約9到10公分的梯形硬塊[38]。
病狀
在2003年,.M. Rothschild等人利用電腦斷層掃描檢查恐龍化石中的腫瘤,共有10,000的化石接受掃描。只有數個鴨嘴龍科發現了腫瘤,包含:埃德蒙頓龍、短冠龍、計氏龍、巴克龍。一頭埃德蒙頓龍的化石則發現了腫瘤,包含血管瘤、韌帶樣纖維瘤、轉移癌、骨母細胞瘤等病狀。這些腫瘤可能是由遺傳規律造成的[39]。
在224塊埃德蒙頓龍的趾骨中,有2.2%發現這個骨軟骨炎等病症,骨軟骨炎是因為軟骨在生長階段未能骨化成骨頭所致。骨軟骨炎的根本原因則未知,目前已有遺傳規律、外傷、攝食強度、血液供應的改變、過量的甲狀腺或荷爾蒙、其他成長因素的缺陷…等理論[40]。
食性
同位素研究
科學家曾分析埃德蒙頓龍牙齒中的琺瑯質,研究其中的碳與氧同位素比例,以得知埃德蒙頓龍的時性與生理特徵。當動物進食、喝水、呼吸時,會吸收大自然中的氧與碳,反應在骨頭組織上。不同的外部與內部因素,會影響氧與碳的同位素,例如:該動物的生理狀況、鹽度、氣候。如果骨頭在化石化與成岩的過程中沒有被影響,科學家可以研究其中的同位素,以瞭解該動物生前的各項因素。以特定植物為食的動物,或是有特定消化過程的動物,身體內會形成特定比例的同位素。形成牙齒琺瑯質的礦物質,使牙齒琺瑯質成為身體中最不受化學變化影響的部分[15]。
在2004年,Kathryn Thomas與Sandra Carlson利用三顆牙齒來研究埃德蒙頓龍的齒系生長狀況,這三顆牙齒發現於南達科他州科森縣的一個屍骨層,屬於海爾河組,分別是幼年、亞成年、成年的牙齒。他們使用微鑽孔技術,挖出牙齒的內部組織,研究其中的氧同位素比例變化。這三顆牙齒的氧同位素比例變化,似乎與溫暖/乾旱、寒冷/潮濕的氣候變化相關聯。Thomas與Carlson認為這代表者季節變化,也可能是埃德蒙頓龍具有遷徙的習性;但遷徙動物會生存在氣候相近、或有特定食物來源的地區,應會造成平均化的氧同位素比例變化。一顆牙齒的生長,短於0.65年,年輕埃德蒙頓龍的牙齒生長較快[15]。
埃德蒙頓龍的碳同位素數值大,以現代哺乳動物而言,代表牠們同時以富含C3的植物(大部分植物),與富含C4的植物(草)為食;但是,白堊紀晚期的富含C4的植物相當稀少。Thomas與Carlson提出可能影響埃德蒙頓龍體內碳同位素的原因,認為這些埃德蒙頓龍最有可能以裸子植物為食,以及西部內陸海道沿岸的耐鹽植物為食;或者是,恐龍與哺乳動物的生理特徵不同,使恐龍的體內形成不同的碳同位素含量。也有可能是以上各因素的綜合結果[15]。
進食方式

如同大部分鴨嘴龍類,埃德蒙頓龍的頭骨類似現代的鴨。頭骨前段平坦、寬廣,具有適合咬斷樹葉、樹枝的喙狀嘴。埃德蒙頓龍具有數千顆牙齒。牠們可以將植物置於兩側的頰部,並以牙齒咀嚼粗糙的食物。這些牙齒以很緊密的方式排列,形成多達數十列齒系(Dental batteries)。新的牙齒會不斷的生長來取代脫落的牙齒。當嘴部咬合時,上頜骨可以向外彎折。食物通常包括針葉樹的針葉、種子及樹枝,可以在埃德蒙頓龍化石的體腔內發現這些植物化石。
在1984年,大衛·威顯穆沛(David B . Weishampel)首次提出鴨嘴龍類的進食方式研究。威顯穆沛根據鴨嘴龍類的頭部構造,提出下頜可以做出前後移動的動作,而當嘴部咬合時,上頜的骨頭會往外側彎曲。當咬合時,上下頜的牙齒會將嘴中的食物磨碎[7][41]。這種動作類似哺乳動物的咀嚼動作,但以不同的方式進行[42]。威顯穆沛研究的重要性在於,根據牙齒的磨痕方向,提出頜部的運動方向。雖然其他方向的運動也有可能造成類似的牙齒磨痕。科學家已利用目前技術檢驗這些可能的其他運動方向[43]。
威顯穆沛當年曾借助電腦來模擬鴨嘴龍類的頭部。在2008年,Natalia Rybczynski等人利用雷射掃描帝王埃德蒙頓龍的頭骨,然後利用更複雜的電腦程式模擬出鴨嘴龍類的頭部。他們發現埃德蒙頓龍咬合時,頭骨之間還有其他次要的運動,而骨頭間的最大分隔約1.3到1.4公分。Rybczynski等人認為威顯穆沛的重建模型有誤,認為有軟組織、齒痕等其他可變因素,需要其他實驗才能重建埃德蒙頓龍咬合時的頭骨動作[42]。Casey Holliday與勞倫斯·威特默(Lawrence Witmer)則發現鳥腳類恐龍的頭部各骨頭間缺乏可動性,與現代有鱗目、鳥類不同。他們推測埃德蒙頓龍的頭骨間關節,其實是軟骨的生長地區[43]。
過去曾有研究宣稱,鴨嘴龍科的一個破碎龍化石曾發現胃石。但該化石是在1900年由巴納姆·布郎(Barnum Brown)發現,被歸類於C. annectens,目前則屬於埃德蒙頓龍的E. annectens。此外,這些胃石其實是化石化過程中,被沖刷、沉積進入的砂礫[44]。
參考資料
- Lambe, Lawrence M. . Memoir 120. Department of Mines, Geological Survey of Canada. 1920: 1–79.
- Gilmore, Charles W. . Bulletin 38. Department of Mines, Geological Survey of Canada. 1924: 13–26.
- Sternberg, Charles M. . Bulletin 44. Department of Mines, Geological Survey of Canada. 1926: 77–84.
- Lull, Richard Swann; and Wright, Nelda E. . Geological Society of America Special Paper 40. Geological Society of America. 1942: 50–93.
- Glut, Donald F. . . Jefferson, North Carolina: McFarland & Co. 1997: 389-396. ISBN 0-89950-917-7.
- Lambert, David; and the Diagram Group. . New York: Avon Books. 1990: 60. ISBN 978-0-380-75896-8.
- Horner, John R.; Weishampel, David B.; and Forster, Catherine A. . Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.) (编). 2nd. Berkeley: University of California Press. 2004: 438-463. ISBN 0-520-24209-2.
- Lull, Richard Swann; and Wright, Nelda E. . Geological Society of America Special Paper 40. Geological Society of America. 1942: 225.
- Lucas, Frederic A. . Smithsonian Miscellaneous Collections. 2004, 45: 317–320.
- Morris, William J. . Contributions in Science. 1970, 193: 1–14.
- Lull, Richard Swann; and Wright, Nelda E. . Geological Society of America Special Paper 40. Geological Society of America. 1942: 151–164.
- Lull, Richard Swann; and Wright, Nelda E. . Geological Society of America Special Paper 40. Geological Society of America. 1942: 110–117.
- Hopson, James A. . Paleobiology. 1975, 1 (1): 21–43.
- Lull, Richard Swann; and Wright, Nelda E. . Geological Society of America Special Paper 40. Geological Society of America. 1942: 128–130.
- Stanton Thomas, Kathryn J.; and Carlson, Sandra J. . Palaeogeography, Palaeoclimatology, and Palaeoecology. 2004, 206 (2004): 257–287. doi:10.1016/j.palaeo.2004.01.007.
- Ostrom, John H. . American Journal of Science: 975–997.
- Galton, Peter M. . Journal of Paleontology. 1970, 44 (3): 464–473.
- Gates, Terry A.; Sampson, Scott D. . Zoological Journal of the Linnean Society. 2007, 151 (2): 351–376. doi:10.1111/j.1096-3642.2007.00349.x.
- Weishampel, David B.; and Horner, Jack R. . Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.) (编). 1st. Berkeley: University of California Press. 1990: 534–561. ISBN 0-520-06727-4.
- Lull, Richard Swann; and Wright, Nelda E. . Geological Society of America Special Paper 40. Geological Society of America. 1942: 48.
- Carpenter, Kenneth. . Gaia. 1998, 15: 135–144 [2008-12-19]. (原始内容存档于2007-11-17). [not printed until 2000]
- Campagna, Tony. . Prehistoric Times. 2000, 40: 18–19.
- Russell, Dale A.; and Chamney, T. P. . National Museum of Canada Natural History Papers. 1967, 35: 1–35.
- . ScienceDaily. ScienceDaily. 2007-07-03 [2008-12-07]. (原始内容存档于2020-11-09).
- Eberth, David A. (PDF). Canadian Society of Petroleum Geology Diamond Jubilee Convention, Programs and Abstracts. 2002, 117: (cd) [2009-01-01]. (原始内容 (pdf)存档于2007-11-13).
- Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loueff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth M.P.; and Noto, Christopher N. . Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.) (编). 2nd. Berkeley: University of California Press. 2004: 517–606. ISBN 0-520-24209-2.
- Wu, X-C.; Brinkman, D.B.; Eberth, D.A.; and Braman, D.R. . Canadian Journal of Earth Science. 2007, 44 (9): 1243–1265.
- Behrensmeyer, Anna K. (Excel spreadsheet). Bonebeds: Genesis, Analysis, and Paleobiological Significance. University of Chicago Press. 2007 [2008-12-07]. (原始内容存档于2019-06-30).
- Chadwick, Arthur; Spencer, Lee, and Turner, Larry. . Journal of Vertebrate Paleontology. 2006, 26 (3, suppl.): 49A.
- Jacobsen,, Aase Roland; and Ryan, Michael J. . Journal of Vertebrate Paleontology. 2000, 19 (3, suppl.): 55A.
- Gould, Rebecca; Larson, Robb; and Nellermoe, Ron. . Journal of Vertebrate Paleontology. 2003, 23 (3, suppl.): 56A–57A.
- Bell, Phil R.; and Snively, E. . Alcheringa. 2008, 32 (3): 271–284. doi:10.1080/03115510802096101.
- Lloyd, Robin. . LiveScience.com. Imaginova. 2008-12-04 [2008-12-11].
- Osborn, Henry Fairfield. . Science. 1909, 29 (750): 793–795.
- Osborn, Henry Fairfield. (pdf (very large; 76,048 kb)). Memoirs of the American Museum of Natural History. 1912, 1: 33–54 [2009-01-12]. (原始内容存档于2014-12-21).
- . National Geographic News. 2007-12-03 [2007-12-03]. (原始内容存档于2012-10-31).
- Lee, Christopher. . Washington Post. 2007-12-03 [2007-12-03]. (原始内容存档于2012-11-03).
- Lyson, Tyler R., Hanks, H. Douglas, and Tremain, Emily S. (2003) "New skin structures from a juvenile Edmontosaurus from the Late Cretaceous of North Dakota (页面存档备份,存于)". Geological Society of America. Last accessed 2008-08-02.
- Rothschild, B.M.; Tanke, D.H., Helbling II, M. and Martin, L.D. . Naturwissenschaften. 2003, 90 (11): 495–500 [2008-07-25]. doi:10.1007/s00114-003-0473-9.
- Rothschild, Bruce; and Tanke, Darren H. . Carpenter, Kenneth (ed.) (编). . Bloomington and Indianapolis: Indiana University Press. 2007: 171–183. ISBN 0-253-34817-X.
- Weishampel, David B. . Advances in Anatomy, Embryology, and Cell Biology 87. Berlin; New York: Springer-Verlag. 1984. ISBN 0387131140. ISSN 0301-5556. PMID 6464809.
- Rybczynski, Natalia; Tirabasso, Alex; Bloskie, Paul; Cuthbertson, Robin; and Holliday, Casey. . Palaeontologia Electronica. 2008, 11 (2): online publication [2008-08-10]. (原始内容存档于2008-10-05).
- Holliday, Casey M.; and Witmer, Lawrence M. . Journal of Vertebrate Paleontology. 2008, 28 (4): 1073–1088.
- Creisler, Benjamin S. . Carpenter, Kenneth (ed.) (编). . Bloomington and Indianapolis: Indiana University Press. 2007: 185–210. ISBN 0-253-34817-X.