弓鲨目
弓鲨目又称弓鲛目(学名:Hybodontiformes),也称为"Hybodonts",是生存于石炭纪至古新世的一类已灭绝的板鳃亚纲(鲨鱼、魟鱼和鳐鱼)。它们构成了与新鲨类最接近的种群,即现代鲨鱼和鳐鱼的姐妹群。 弓鲛目根据它们的锥形齿来命名并区分。它们是侏罗纪时期欧洲和北美鲨鱼的主要群体,该目大部分种类灭绝于晚白垩世,可能是由于在与其他鲨鱼的竞争中处于劣势,不过皱齿鲨属等一直存活到了古新世[1]。矛鲛属是最后的弓鲛目之一,它独特的锯齿状细刺与最后的非鸟恐龙的化石一起出现在怀俄明州的淡水沉积物中,包括霸王龙和三角龙。弓鲛目物种在化石记录中主要根据牙齿和鳍刺的不同进行鉴定。众所周知,它们在淡水和咸水环境中均有分布。
| 弓鮫目 化石时期: | |
|---|---|
 | |
| 莫氏皱齿鲨,一种体长可达7米的晚白垩世鲨鱼,其是否属于弓鲨目存在争议 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 软骨鱼纲 Chondrichthyes |
| 亚纲: | 板鳃亚纲 Elasmobranchii |
| 下纲: | 真鲨下纲 Euselachii |
| 目: | †弓鮫目 Hybodontiformes Owen, 1846 |
| 下属分类 | |
词源
"hybodont"来自希腊文ὕβος或ὑβός(意为驼峰或驼背)和andδούς(δοντ,意为牙齿),该名称是根据其圆锥形的牙齿命名的。
系统发育
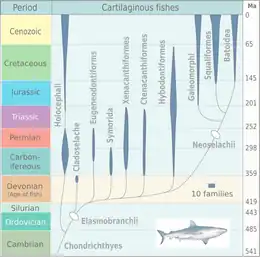
弓鲛目是新鲨类外的一类软骨鱼,与栉棘鲨目和新鲨类一起被归为鲨鳐下纲。 弓鲛目目前有无尖齿鱼科、弓鲛科、矛鲛科、多尖齿鱼科、翼柱头鲨科这五个科,其中滑齿鲨属是多尖齿鱼科的代表,弓鲛属是弓鲛科的代表,而蒙氏翼柱头鲨是翼柱头鲨科的代表[3]。
地质记录
起源
弓鲛目最早是路易士·阿格西在1837年基于孤立的化石牙齿进行描述的。最早的弓鲛目化石来自石炭纪,包括三锋鲛属和其他来自苏格兰方解石与砂岩的鱼类[4]1911年,齐特尔(Zittel)首先将弓鲛目与现生鲨鱼区分开[5]。
尽管最早的弓鲛目化石是石炭纪的,但在泥盆纪早期它们可能已经从与新鲨类(现代鲨鱼)的共同祖先中分化出来了[6]。
二叠纪
现在称为弓鲛目的属包括许多种,例如弓鲛属、无尖齿鱼属、星棘鲨属、矛鲛属和滑齿鲨属。从阿曼的二叠纪矿床中回收了海牙样本,表明弓鲛目在二叠纪期间生活在新特提斯洋[7]。
这项研究与来自得克萨斯州的其他研究相结合,表明在二叠纪期间,弓鲛目的发展已经很成熟,并且在某些地方占主导地位[7]。总的来说,二叠纪弓鲛目的化石记录是有限的。最初的假设是,二叠纪末期灭绝并没有显着影响弓鲛目的多样性,与此相反,人们后来认为二叠纪-三叠纪灭绝事件之前的五千万年,二叠纪弓鲛目的多样性有所下降[7]。但是,最近在阿曼发现的样本表明,二叠纪的弓鲛目的多样性一直延伸到二叠纪末期[7]。
三叠纪-早侏罗世
来自加拿大艾伯塔省硫山地层的三叠纪下层Vega-Phroso粉砂岩中发现了保存完好的Wapitiodus aplopagus化石标本,它们在灭绝中幸存下来,并且在三叠纪早期数量很多[8]。
三叠纪期间弓鲛目的多样性达到了巅峰,在三叠纪和早侏罗世,无论是在海洋环境中还是非海洋环境中,弓鲛目都是最主要的鲨鱼种群[9]。一项对英格兰中侏罗世化石的研究分析了11个属的20个物种,表明当时的弓鲛目在蓬勃发展[9]。
栖息地
.jpg.webp)
弓鲛目化石在海相到河流相(河流沉积)的沉积环境中被发现。当它们第一次进化时,它们生活在海洋和淡水系统中。虽然多数弓鲛属生活在淡水中,但在泻湖和其他封闭的沉积环境中发现的中侏罗世样品中有一个弓鲛属进入更为狭小的环境的例子[9]。在泰国淡水Khorat群(上侏罗世-阿普第阶(下白垩纪))中发现的7枚弓鲛属的牙齿被描述。
形态与牙齿
通常根据牙齿和鳍脊骨化石的大小和形状来描述和鉴定弓鲛目物种。弓鲛目畸形的牙齿被认为具有突出的齿尖,该齿尖高于侧齿[14]。弓鲛的种类通常是根据它们的牙齿化石确定的,这些牙齿化石比鲨鱼骨骼的其余部分更有可能保存在岩石记录中。化石记录中的牙齿很丰富,因为鲨鱼一生中都会更换它们,并且牙齿具有抗腐蚀能力。由于牙齿的根部没有很好地附着在牙冠上,因此弓鲛的牙齿通常被保存为不完整的化石[14]。根据牙齿的形状,最初将它们分为两类[15]。一组牙齿的齿尖尖而没有牙髓腔,这些被称为骨牙;另一组具有不同的齿尖排列,并具有牙髓腔,这些被称为正牙[16]。例如,弓鲛目物种Heterophychodus steinmanni的牙齿上具有垂直排列的牙本质血管,也称为“管状牙本质”[10]。这些骨质疏松牙齿的冠部覆盖有单层的同种生物。
弓鲛目的特征是有两个背鳍,每个背鳍之前都有特定形状的棘刺。鳍状棘刺的形状用于区分其他鲨鱼种群和不同种类的弓鲛目[4]。鳍棘细长,并朝着动物的背部轻轻弯曲。雄性弓鲛目物种的头部有小刺。弓鲛目的下颚粗大而厚实,根据饮食和牙齿的不同而有所不同[17]。弓鲛目有成对的鳍,用于转向,尾鳍完全异种,或尾部不平整,背鳍比腹鳍更延伸。
最完整的弓鲛目化石之一是巴西东北部桑塔纳组(早白垩世)的Tribodus样本[18]。回收了一个三维标本,揭示了胸鳍的形状以及它们如何附着在肩骨上。
行为
弓鲛目可能是游速较慢的鲨鱼,并使用其成对的鳍进行操纵和稳定。 弓鲛属是一类典型的弓鲛目,被认为游速缓慢,但偶尔会突然爆发,使其成为快速移动追捕猎物的活跃掠食者[11]。牙弓有各种各样的牙齿形状,这种多样性表明它们利用了多种食物来源[14]。有人认为,某些齿宽较宽,更扁平的舌齿专门用于粉碎硬壳猎物。 滑齿鲨属牙齿上发达的磨损小面表明某些齿状牙齿压碎了它们的食物[9]。泰国描述的物种具有多种牙齿形状,表明有多种进食习惯。球形牙齿被用来压碎具有硬壳的底栖猎物[10]。其他一些则是机会主义者,其饮食中有大量的软体动物[19]。关于弓鲛目的生殖习性知之甚少,一项研究在吉尔吉斯斯坦的三叠纪淡水湖沉积物中发现了丰富的牙齿化石和卵袋[19]。基于丰富的卵和幼年个体化石以及数量有限的成年个体标本,该遗址被解释为一个古老的鲨鱼繁殖地[19]。
参考资料
- The paleobioloy Database Ptychodontidae entry (页面存档备份,存于) accessed on 8/23/09
- Benton, M. J. (2005) Vertebrate Palaeontology, Blackwell, 3rd edition, Fig 7.13 on page 185.
- Eaton, J. G., Cifelli, R. L., Hutchison, J. H., Kirkland, J. I., and Parrish, M. J., 1999, Cretaceous vertebrate faunas from the Kaiparowits Plateau, south-central Utah: Utah Geological Survey Miscellaneous Publication, v. 99-1.
- Maisey, J. G., 1978, Growth and form of spines in hybodont sharks: Palaeontology, v. 21, no. 3, p. 657-666.
- Zittel, K. von, 1911, Grunzuege der Palaontologie, 2 ed. II. Abt. Vertebrata, vii + 598 pp. R. Oldenburg。Verlag, Muchen, Berlin.
- Coates, M. I.,and Gess, R. W., 2007,A new sreconstruction of Onychoselache.Traquairi, comments on early Chondrichthyan pectoralgirdles and hybodontiform phylogeny: Palaeontology, v. Devonian50, (no. 6,)p. 1421-1446.
- Koot, M. B., Cuny, G., Tintori, A., and Twitchett, R. J., 2013, A new diverse shark fauna from the Wordian (Middle Permian) Khuff Formation in the interior Haushi-Huqf area, Sultanate of Oman: Palaeontology, v. 56, no. 2, p. 303-343.
- Mutter, etR. alJ., De Blanger, K., and Neuman, A. G., 2007, Elasmobranchs from the Lower Triassic Sulphur Mountain Formation near Wapiti Lake (BC, Canada): Zoological Journal of the Linnean Society, v. 149, no. 3, p. 309-337.
- Rees, J. A. N., and Underwood, C. J., 2008, Hybodont sharks of the English Bathonian and Callovian (Middle Jurassic): Palaeontology, v. 51, no. 1, p. 117-147.
- Cuny, G., Suteethorn, V., Buffetaut, E., and Philippe, M., 2003, Hybodont sharks from the Mesozoic Khorat Group of Thailand: Mahasarakham University Journal, v. 22.
- Maisey, J. G., 2012, What is an ‘elasmobranch’? The impact of palaeontology in understanding elasmobranch phylogeny and evolution: Journal of Fish Biology, v. 80, no. 5, p. 918-951.
- Everhart, Michael; Caggiano, Tom. 4 (4): 125–136.
- Hoffman, Brian. . Journal of Paleontology. July 2016, 90 (4): 741–762. S2CID 132252846. doi:10.1017/jpa.2016.64.
- Koot, M. B., Cuny, G., Tintori, A., and Twitchett, R. J., 2013, A new diverse shark fauna from the Wordian (Middle Permian) Khuff Formation in the interior Haushi-Huqf area, Sultanate of Oman: Palaeontology, v. 56, no. 2, p. 303-343.
- Agassiz, L., 1833-1844, Recherches sur les poisons fossils. Neuchatel, 5 vols. 1420 pp. with supplement.
- Maisey, J. G., 1982, The anatomy and interrelationships of Mesozoic hybodont sharks: American Museum Novitates, v. 2724.
- Carvalho, Marcelo, http://www.amnh.org/learn/sharks/Resource1 (页面存档备份,存于)
- Lane, J. A., and Maisey, J. G., 2009, Pectoral Anatomy of Tribodus limae (Elasmobranchii: Hybodontiformes) from the Lower Cretaceous of Northeastern Brazil: Journal of Vertebrate Paleontology, v. 29, no. 1, p. 25-38.
- Fischer, J. A. N., Voigt, S., Schneider, J. W., Buchwitz, M., and Voigt, S., 2011,A selachian freshwater fauna from the Triassic of Kyrgyzstan and its implication for Mesozoic shark nurseries: Journal of Vertebrate Paleontology, v. 31, no. 5, p. 937-953.