细胞毒性T细胞
细胞毒性T细胞(英語:,TC或CTL,简称胞毒T細胞)又称杀手T细胞()、TC细胞、胞殺T細胞、效應T細胞或CD8+ T細胞,屬於T細胞的一種,可以殺死癌細胞、受病毒感染的細胞,以及其他受損細胞。胞毒T细胞属于后天免疫系統的成员,当其杀伤活性开始表现时,属于先天免疫系统的NK细胞活性就会逐步降低。T細胞的一種。 執行清除被感染細胞。 分泌穿孔素使被感染細胞穿孔,導致細胞凋亡。
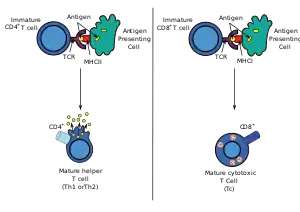
大多数细胞毒性T细胞会表达T細胞受體(TCR),此受體可以识别特定的抗原,而那些抗原常常是由癌細胞和病毒製造的。胞內抗原會與第一型MHC分子結合,並被帶到細胞表面,以利於T細胞辨識。T細胞一旦透過T細胞受體辨識出特定的抗原,便會摧毀該細胞。
有些T細胞受體中含有一種稱為CD8的醣蛋白,可與MHC1類分子結合,故這些細胞又名CD8+ T細胞。
和第一型MHC分子結合的CD8 T細胞會發育為細胞毒性T細胞,而CD8分子則會持續牢牢地抓著第一型MHC分子,不讓目標細胞離開,以進行細胞毒殺。
發育

免疫系統必須有能力辨識數百萬種抗原,但身體內卻只有30,000對基因,所以不可能一個抗原就花費一組基因來辨識。因此,身體內採用的勢必是另一套機制。骨髓中的未成熟白血球DNA會改變,製造出特定的白血球受體,而每一種白血球則可以與不同的抗原結合。但這種方式製造出來的白血球有可能會與身體中的物質結合,如果放任這些白血球四處發生免疫反應,稱為自體免疫疾病,嚴重者可致死。因此,身體將這些未成熟的白血球送到胸腺,將這些會與體內物質結合的白血球摧毀。
T細胞受體(T cell receptor,簡稱TCRs)有兩個部分,分別是α鏈和β鏈(有些有γ鏈和δ鏈)。骨髓中的造血幹細胞會移動到胸腺,並在胸腺進行β鏈的VDJ重組,稱為前T細胞受體。如果重組成功,那麼原先的α鏈TCR DNA就會轉為αβ TCR複合體。這種高變異的重組模式,可以產生數百萬種不同的T細胞,可以辨識出不同的抗原。大部分的T細胞的受體為αβ TCR(αβ T細胞)但有一些表皮中的T細胞含有γδ TCR(γδ T細胞),可以辨識一些非蛋白質性抗原。
T細胞受體穩定的幼年T細胞會同時表現CD4和CD8協同受體,因此被稱為雙陽性T細胞(double-positive T cell,簡稱DP T cell),胸腺中的雙陽性T細胞會接觸到大量的自身抗原,並進行兩種株落選擇:
經過這兩道篩檢之後所剩下來的T細胞才會成熟,會轉變為單陽性T細胞(CD4+或CD8+),如果是CD8+,那麼成熟的T細胞就會辨識第一類MHC分子;如果是CD4+,那麼成熟的T細胞就會辨識第二類MHC分子。決定辨識CD8的T細胞就會變成細胞毒性T細胞。
活化
除了一些少數細胞和沒有細胞核的細胞(如紅血球),宿主的細胞表面幾乎都會表現第一類MHC分子。當細胞被病毒感染時(或其他胞內病原體),細胞會降解外來蛋白質,並由第一類MHC分子將蛋白質片段表現於細胞表面,以利於CD8+ T細胞辨識,此動作稱為抗原呈現。
細胞毒性T細胞的活化取決於T細胞表面的分子和抗原呈递细胞表面分子之間的交互作用。例如雙訊息模型:
| 訊息 | T細胞 | APC | 描述 |
| 第一訊息 | TCR | 第一類MHC分子 | CD8輔助受體和第一類MHC分子之間會引發交互作用以穩定信息分子。 |
| 第二訊息 | T細胞上的CD8 | CD80或CD86(又稱B7-1和B7-2) | 由輔助型T細胞釋出,可活化細胞毒性T細胞。CD80和CD86在T細胞活化中扮演著「协同刺激信号」的角色。 |
殺手T細胞的活化大多是靠TCR偵測到抗原所引發,但也有少部分是靠替代路徑。舉例來說,研究顯示細胞毒性T細胞如果被其他CD8 T細胞視為目標,會引起對於後者的耐受作用。[3]
細胞毒性T細胞一旦活化,便會在IL-2的刺激下開始快速增殖。IL-2是一種T細胞的成長和细胞分化因子,使T細胞能夠更有效的清除體內含有抗原的体细胞。
作用機制
當細胞毒性T細胞遇到受感染或是不健康的體細胞,細胞毒性T細胞會釋放出細胞毒素,如穿孔素、顆粒酶,以及顆粒溶解素。這些酵素會進入目標細胞的細胞質,並使細胞內的丝氨酸蛋白酶誘發胱天蛋白酶的級聯反應,這個反應會使一連串的胱胺酸蛋白酶活化,引發细胞凋亡。
另外一種誘發細胞凋亡的途徑是藉由細胞表面受體的結合。細胞毒性T細胞表面上有種表面蛋白,叫FAS配體(FasL,又稱Apo1L、CD95L)。這種蛋白會和目標細胞上的Fas受體(Apo1,又稱CD95)結合[4]。Fas與FasL結合後,會引來死亡誘導訊號複合體(death-induced signaling complex,簡稱DISC),Fas關聯死亡區域(Fas-associated death domain,FADD)會隨著DISC移位,引來胱天蛋白酶原8和10[5],之後胱天蛋白酶會被胱天蛋白酶3、6,和7活化,導致如lamin A、lamin B1、lamin B2、PARP(聚ADP核糖聚合酶),和DNAPK(DNA活化蛋白激酶)等蛋白被水解,導致細胞凋亡。
疾病病理學
在B型肝炎病毒(HBV)的感染上,細胞毒性T細胞扮演了一個相當重要的致病原。因為被HBV感染的肝細胞會被細胞毒性T細胞所毒殺,因此造成肝部的嚴重損壞。[6]近期研究顯示,血小板可吸引專門對付病毒的細胞毒性T細胞來到受感染的肝臟。[7]
參見
參考文獻
- S. Munir Alam, Paul J. Travers, Jay L. Wung, Wade Nasholds, Stella Redpath, Stephen C. Jameson, Nicholas R. J. Gascoigne. . Nature. 1996/06, 381 (6583): 616–620 [2018-04-02]. ISSN 1476-4687. doi:10.1038/381616a0. (原始内容存档于2010-06-26) (英语).
- Timothy K. Starr, Stephen C. Jameson, Kristin A. Hogquist. . Annual Review of Immunology. 2003-04-01, 21 (1): 139–176 [2018-04-02]. ISSN 0732-0582. doi:10.1146/annurev.immunol.21.120601.141107. (原始内容存档于2022-04-12).
- Milstein, O., Hagin, D., Lask, A., Reich-Zeliger, S., Shezan E., Ophir E., Eidelshtein Y., Afik R., Antebi YE., Dustin ML. and Reisner Y. (2011) CTLs respond with activation and granule secretion when serving target for T cell recognition. Blood 117,1042-1052 (页面存档备份,存于)
- Andersen, MH; Schrama, D; Thor Straten, P; Becker, JC. . J. Invest. Dermatol. 2006, 126 (1): 32–41. PMID 16417215. doi:10.1038/sj.jid.5700001. 已忽略未知参数
|author-name-separator=(帮助); 已忽略未知参数|author-separator=(帮助) - M. Muzio, A. M. Chinnaiyan, F. C. Kischkel, K. O'Rourke, A. Shevchenko, J. Ni, C. Scaffidi, J. D. Bretz, M. Zhang, R. Gentz, M. Mann, P. H. Krammer, M. E. Peter, V. M. Dixit. . Cell. 1996-06-14, 85 (6): 817–827 [2019-02-13]. ISSN 0092-8674. PMID 8681377. (原始内容存档于2019-06-03).
- Iannacone, Matteo; Sitia, Giovanni; Guidotti, Luca G. . Future Virology. 2006, 1 (2): 189–96. doi:10.2217/17460794.1.2.189.
- Iannacone, Matteo; Sitia, Giovanni; Isogawa, Masanori; Marchese, Patrizia; Castro, Maria G; Lowenstein, Pedro R; Chisari, Francis V; Ruggeri, Zaverio M; Guidotti, Luca G. . Nature Medicine. 2005, 11 (11): 1167–9. PMC 2908083
 . PMID 16258538. doi:10.1038/nm1317.
. PMID 16258538. doi:10.1038/nm1317. - Subramanian S and Ramalingam K. . Asian Journal of Microbiology, Biotechnology and Environmental Sciences. 2005, 7 (2): 227–233. ISSN 0972-3005.
外部連結
- (英文)T-cell Group - Cardiff University
- (英文)Malaria (Flash Animation)