环醇假说
环醇假说(cyclol hypothesis),為第一个折叠的球状蛋白质结构模型概念[1] ,这是多萝西·林奇在20世纪30年代基于三个假设提出的。首先,该假设假设两个肽键由Cyclol反应交联而成(图1),这些交联键是共价的肽的同系物之间的氢键,而非“共价键”。这些反应已被发现存在于麦角肽和其他化合物中。第二,该假设提出在某些条件下,氨基酸会自然地在最大数量上进行Cyclol交联反应,生成Cyclol分子(图2)和Cyclol结构(图3)。这些Cyclol分子和Cyclol结构从来没有被观察到。最后,该假设认为球状蛋白质的三级结构对应的正多面體和半规则的多面体形成的Cyclol结构是没有自由边缘的。这种“封闭式的Cyclol”分子也没有被观察到。
尽管后来的数据表明,这种本源的球状蛋白质结构需要被修正,几种元素的Cyclol模型也得到了验证,比如Cyclol反应本身和该假设,即疏水性相互作用主要负责蛋白质的折叠。Cyclol假设是一个更精确的DNA双螺旋结构和蛋白质二级结构模型假设,同时激励许许多多的科学家研究蛋白质结构和化学性质的相关问题。Cyclol假设的提出和测试也为经验证伪性作为科学方法的一部分提供了一个很好的例证。
历史背景
20世纪30年代中期,由西奥多·斯维德伯格提出的超高速离心研究表明蛋白质有一个明确的化学结构,并没有聚合的小分子 [2]。 同样的研究似乎表明,蛋白质的分子量与几个有明确定义的整数有一定的关系[3], 例如Mw = 2p3qDa,其中p和q都是非负整数[4]。 然而,确定确切的分子量和蛋白质分子中的氨基酸数量却相当困难。斯维德伯格还表示,溶液条件下的变化可能导致蛋白质分解成许多子单元,即现在已知的四级机构变化[5]。
在那时,蛋白质的化学结构仍未确定[6]。 最被大众接受(并最终被证实正确)的蛋白质结构假设指出蛋白质是线性的多肽,即无支链聚合物的氨基酸以肽键相连[7][8]。 然而,一个典型的蛋白质是非常长—数百个氨基酸基团—几位著名的科学家不能确定这样的长的线性大分子是否能稳定的存在于溶液中[9][10]。 进一步怀疑产生蛋白质的多肽性质是因为观察到某些酶分解蛋白质而不是多肽,另一些分解多肽但是没有折叠蛋白质[11]。 尝试在试管中合成蛋白质是不成功的,这主要是因为氨基酸的手性;天然存在的蛋白质是只由左旋的氨基酸组成的。因此,非正统的蛋白质结构的化学模型也被列入考虑范围,如埃米尔·阿布德哈尔登的二酮哌嗪假设 [12][13]。 但是,没有一种非正统的模型能解释为什么蛋白质只产生氨基酸和多肽、蛋白质水解产物。由Linderstom-Lang [14]澄清,这些蛋白质水解数据显示,变性的蛋白质就是多肽,不过没有数据显示蛋白质的折叠结构;因此,变性可能涉及一个将折叠的蛋白质转化为多肽的化学变化。
1910年,蛋白质的变性过程(区别于凝固)被哈丽叶和查尔斯·马丁[15]发现, 但其本质仍然是个迷。提姆·安森和阿尔弗雷德·米尔斯基表示变性是一个可逆的,共存的过程[16], 致使许多化学基团能参与化学反应,包括裂解酶[16][17]. 1929年,吴宪正确地假设变性相当于蛋白质打开,换句话说也就是一个纯粹的导致氨基酸侧链暴露在溶剂中的构象变化.[18]. 吴的假设也在1936年分别被米尔斯基和萊納斯·鮑林进一步完善[19]. 然而,研究蛋白质的科学家不能排除变性是一个蛋白质结构化学变化的可能性[17],该假设知道20世纪50年代才被认为是有一定(相对较小的)可能性[20][21].
1911年,X射线晶体学才开始成为一门学科,同时相对迅速地推进了从简单的盐晶到复杂分子的晶体,比如胆固醇。然而,即使是最小的蛋白质也具有超过1000个原子,这使得他们的结构更加的复杂。虽然胰岛素和其他蛋白质的结构直到20世纪60年代末才得以解决,但是多萝西·克劳福特·霍奇金早在1934年就已采集小分子蛋白质,也即胰岛素的晶体结构数据。然而,20世纪30年代初许多天然纤维,比如羊毛和头发的开创性的X射线纤维衍射数据就已经被威廉·阿斯特伯里收集。他提出了二级结构元素的基础模型,如蛋白质的α螺旋和β折叠。
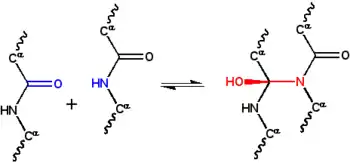
20世纪30年代,由于人们对于蛋白质的结构知之甚少,对物质结构起到物理稳定作用也同样未知。威廉 阿斯特伯里推测是β折叠中的氢键对纤维蛋白的结构起到稳定作用[22][23]. 1932年,多萝西·乔丹·劳埃德也提出了氢键稳定球状蛋白结构的想法[24][25], 后来也得到阿尔弗雷德·米尔斯基和莱纳斯·鲍林的支持[19] 在牛津初级科学协会1933年阿斯特伯里的讲座当中,物理学家弗雷德里克·弗兰克提出纤维蛋白α—角蛋白可能是由一种替代机制稳定,也就是说是由上述的Cyclol反应共价交联形成肽键。[26], Cyclol交联将两个肽基紧密结合在一起;N原子和C原子相距~1.5 Å ,而在一个典型的氢键当中相距~3 Å。J·D·博纳尔对这个想法很感兴趣,他向数学家多萝西·林奇表示,这个想法可能有助于了解蛋白质的结构。
基本理论
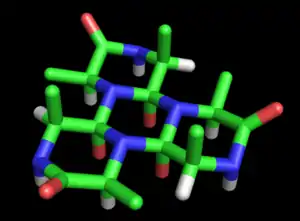
林奇将这个想法发展成一个完整的蛋白质结构模型。在她的第一篇论文(1936年)中奠定了基础的Cyclol模型[27]. 她指出,多肽环化形成闭合环状结构(真实存在),这些环状结构通过Cyclol反应内部交联(也真实存在,但是非常少见)。假设肽键的Cyclol构型比酰胺键更加稳定,林奇得出这样的结论:特定的环状肽会产生最大量的Cyclol构型(比如Cyclol 6,图2)。如果化学键被视为具有相同的长度,那么这种Cyclol分子将具有六边形对称性,大约为1.5 Å;相对而言N-C键C-C键的长度分别为1.42 Å和1.54 Å。
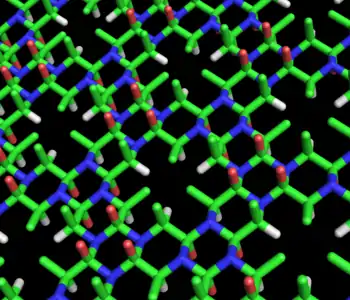
这种环状结构可以无限延长形成Cyclol结构(图3)。这种结构表现出一种远程、准晶体的规律,在林奇认为这很可能存在于蛋白质中,由于它们必须将数以百计的基团紧密的聚合在一起。这种分子和结构的另一个有趣的特点就是它们的氨基酸侧链只从一面沿轴方向向上伸展,而另一面并没有侧链。因此,一个侧面就是一个多肽的完全独立的一句序列,林奇推测这样的结构也许能解释蛋白质结构的独立特性。
在她的第一篇文章中,林奇表示,Cyclol模型只不过是一个假设,蛋白质结构的一个可能有效模型,那么就不得不取得验证。她在这篇文章和后续的文章中,想提出一个明确、可检验的模型,用以验证假设的结论,也用来对通过实验方法取得的结果进行推测。这些目标,她都成功的实现了;然而,实验和进一步的建模显示Cyclol假设对于球状蛋白质是站不住脚的。
能量稳定
在两份同时发给主编的信中(1936年)[28][29],林奇和弗兰克想主编提出关于肽键的Cyclol结构是否在事实上比酰胺键更加稳定的问题。一个相对简单的计算表明,Cyclol结构明显没有酰胺键稳定。因此,除非能确定一个补偿能源确切来源,否则Cyclol模型将被放弃。起初,弗兰克提出,可能会有更好的与周围环境相互作用用以稳定Cyclol结构;后来,林奇和歐文·朗繆爾推测疏水缔合的非极性侧链提供稳定的能量来克服Cyclol反应的能量消耗[30][31] 。
由于Cyclol键的不稳定性为变性特性提供了一个自然解释,因此被视为该模型的一个优势;Cyclol键的逆转 从而形成更加稳定的酰胺结构使整个结构变为一个开放构型,并使其能够与蛋白酶反应,这与实验结果是一致的[32][33]。早起研究表明,蛋白质由于压力造成与由于高温度造成的变性往往处于不同的状态,这被视为支持Cyclol模型可能性的理论.[34]。
朗缪尔—林奇疏水性稳定假设和Cyclol模型的衰落,主要是由于莱纳斯·鲍林的影响,他提出了氢键使蛋白质结构稳定的假设。20年之后疏水性相互作用被确认为是蛋白质折叠的主要动力[35]。
位阻互补
在她的第三篇论文(1936年)[36]中,林奇指出,许多“具有生理活性”的物质,比如,胆固醇,由稠合的碳六元环组成,因此,可能成为Cyclol分子没有氨基酸侧链一侧的立体互补构型。林奇提出,立体互补是判定一个小分子是否能够结合蛋白质的主要因素之一。 林奇推测,蛋白质负责所有生物分子的合成。注意到细胞只有在极度饥饿的情况下才会消化蛋白质,林奇进一步推测,生物生存离不开蛋白质。
混合模式
从一开始,Cyclol反应被认为氢键的共价键模拟形式。因此,人们很自然地考虑这两种键的混合模型。这是林奇第四篇论文(1936年)关于Cyclol模型的主题[37],多萝西·乔丹·劳埃德也参与了该论文的撰写,是她首先提出了氢键稳定球状蛋白质.[24] 一篇写于1937年的后续的论文中引用其他研究的蛋白质氢键的研究人员,比如莫里斯·罗亚尔·哈金斯和莱纳斯·鲍林[38].
林奇和威廉·阿斯特伯里也写了一篇论文,指出>CαHα和酰胺羰基>C=O酮烯醇异构化的可能性,产生>Cα-C(OHα)< 交联结构再次将氧转化为羟基[39]. 这样的反应可能会产生五元环,而经典的Cyclol假设会产生六元环。这种酮烯醇异构化并没有进一步发展[40].
封闭空间结构
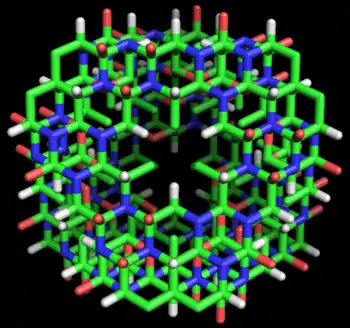
在她关于Cyclol的第五篇论文(1937年)[41]中,林奇确定Cyclol结构可能介入两个平面之间从而形成带有角度的化学键角的条件。她确定了一个简洁的数学模型,其中非平面的六元环的原子可以通过由化学键中点连接而成平面“中心六边形”表示。如果两个平面之间的二面角平分四面体键角即δ= arccos(-1/3)≈109.47°这种“中心六边形”模型使Cyclol结构平面准确介入的情形就非常容易被观察到。
各种各样的封闭的多面体都满足这一标准,其中最简单的是截顶四面体,截顶八面体和八面体,这些都是柏拉图式固体或者半规则的多面体。考虑到第一系列的“封闭式Cyclol结构s”(那些截顶四面体模型),林奇表示,它们的氨基酸数量以72n2 形式增加,其中n 为封闭式Cyclol结构Cn的指数。因此,C1 Cyclol拥有72个基团,C2 Cyclol拥有288个基团,等等。这个推测初步的实验支持来自于麦克斯·伯格曼和卡尔·尼曼 [4],他们对于氨基酸的分析表明,蛋白质由288个氨基酸基团构成 (n=2) 。一般来说,球状蛋白质的Cyclol模型能解释西奥多·斯维德伯格早期的超速离心分析法的结果,这表明该模型使蛋白质的分子量能被几个有明确定义的整数确定[2][3].
Cyclol模型与导致蛋白质折叠的一般特性相一致[42]. (1)离心法研究表明,折叠的蛋白质明显比水更加致密 (~1.4 g/mL),因此,显得更加紧凑;林奇假设说,密实填充意味着规整填料。(2)尽管尺寸偏大,一些蛋白质迅速结晶形成对称晶体,与对称结构的设想是相一致的。(3)蛋白质结合金属离子;由于金属结合位点具有特定的键的几何构型(例如,八面体),这便可以使我们得出一个合理的假设,整个蛋白质也具有相同的晶体几何。(4)如上所述,Cyclol模型为变性和用蛋白酶分解折叠蛋白质的难度提供了一个简单的化学解释。(5)蛋白质被假设认为是负责所有生物分子的合成,也包括其他蛋白质。林奇注意到,一个固定的,统一的结构是有利于蛋白质模板的合成,类似于沃森—弗朗西斯·克里克的DNA自我复制理念。由于许多生物分子,比如由于许多生物分子如碳水化合物和甾醇具有六边形的结构,所以我们可以合理假设,其合成的蛋白质同样具有六边形结构。在她的三篇综述论文中,林奇总结了她的模型及其相对应的分子量实验数据[43].
推测的蛋白质结构
由于提出了一个球状蛋白质的模型,林奇对该模型是否与可用的结构数据相一致进行了调查研究。她推测,牛结核菌色蛋白(523)是一个由72个基团构成的 C1 封闭式Cyclol结构结构[44], 而胃蛋白酶是一个由288个基团构成的C2 封闭式Cyclol结构结构[45][46]。 由于可用于计算蛋白质质量的方法并不准确,因此很难核实这些基团数量的推测是否正确,例如超速离心分析法和化学方法。
林奇还推测,胰岛素是由288个氨基酸基团构成的C2封闭式Cyclol结构结构。有限的胰岛素X射线晶体学数据在林奇认为是能够“证实”她的模型的[47]。 然而,这种解释被严厉地指责为一种草率的结论[48]。 多萝西·克劳福特·霍奇金通过对胰岛素帕特森图的仔细研究,发现它们与Cyclol模型是基本一致的;然而,这种一致性并没有充分地表明Cyclol模型是成立的[49].
衰落
有几个原因证明Cyclol结构是不太可能的。汉斯·纽拉特和亨利·布尔表示Cyclol结构侧链的密实填充与实验所测得的蛋白质膜的密度不相符合[50]。穆利斯·哈金斯计算得到这样的结论:Cyclol结构的几个非键合原子之间的距离比其所允许的范德华半径更小;比如结构内的Hα原子和Cα原子之间的空隙仅仅为1.68 Å(图5)[51]。豪若威兹用化学方法表示蛋白质外侧不会存在很大数量的羟基基团,而这恰恰是Cyclol模型的关键推测[52],梅耶和Hohenemser表示,氨基酸的Cyclol缩合是不存在的,即使作为一个过渡态[53]。伯格曼、尼曼[54]和纽伯格提供了更多的反对Cyclol模型的化学论据[55][56]。红外光谱数据显示蛋白质水解时其中羟基数量并没有改变[57],完整的,折叠蛋白质具有一个完整的酰胺羟基[58]。这两个观察资料否定了Cyclol假设,即在折叠蛋白质中羰基是转化为羟基的。最后,蛋白质含有大量的脯氨酸(通常为5%);由于脯氨酸缺少酰胺氢,脯氨酸中的氮已经形成三个共价键,脯氨酸似乎不能发生Cyclol反应并形成Cyclol结构。鲍林和尼曼给出了一个反对Cyclol模型完善的化学和结构证据概述[59]。此外,1939年一项支持的证据—所有的蛋白质包含288个氨基酸基团的整数倍[4]——被证明是同样不正确的[60]。
林奇回答批评Cyclol模型的空间位阻,自由能,化学品性和剩余基团评论文章。他指出,键角和键长的细微变形减小空间位阻效应,或者至少降低到一个合理的程度[61]。她注意到,在一个分子当中非键合的基团之间的距离小于预计的范德华半径,例如,六甲基苯中甲基原子团之间的距离只有2.93 Å 。关于Cyclol反映的自由能,林奇并不同意鲍林由计算所得的结论,她表示,仅在鲜为人知的分子内能量基础上排除Cyclol模型是不对的[61]。在林奇对化学评论文章的回复中,她表示Cyclol模型不涉及模型化合物和和简单的双分子反应的研究,空间位阻有可能阻止表面羟基反应[62]。关于剩余基团的质疑,林奇在其他剩余基团的数量方面进行了扩展。特别值得一提的是,她提出了一个只由48个基团构成“最小的”封闭式Cyclol结构[63],在这种(不正确的)基础上,可能是第一个提出胰岛素单体的分子量大约为6000 Da[64][65]。
因此,她认为球状蛋白的Cyclol模型是仍然可行的[66][67],甚至提出Cyclol构型是一个细胞骨架组成成分[68]。然而,大多数研究蛋白质的科学家不再相信这种假设,林奇也把她的科研方向转移到X射线晶体学中的数学问题,这在各方向她做出了卓著的贡献。林奇在史密斯学院的同事物理学家格拉迪斯·安斯洛是一个例外,他在20世纪40年代致力于蛋白质和多肽的紫外吸收光谱研究,并在她的结果中解释Cyclol的可能性[69][70]。由于胰岛素的序列开始被弗雷德里克·桑格确认,安斯洛发表了一个带支链的Cyclol三维模型[71],是以林奇1948年提出的“最小Cyclol”模型为基础的[63]。
部分赎回
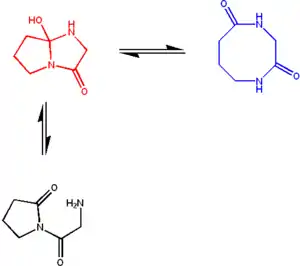
所有Cyclol模型的衰落逐渐导致其自身元素的排斥;一个值得注意的特例就是J·D·贝尔纳对于朗缪尔—林奇蛋白质折叠是由于疏水缔合驱动的假设的短期验收[73]。 然而,20世纪50年代Cyclol键存在于自然生成的小分子环状肽中。
现代术语学的说明是恰当的。经典的Cyclol反应是一个肽基的N-H胺键嵌入另一个基团的C=O羰基;所的化合物现在被称为azacyclol。通过推测,一个羟基基团加入肽基羰基基团后形成一个oxacyclol。同样的,一个硫醇基团加入肽基羰基基团后形成一个thiacyclol [74]。
真菌麦角菌中的oxacyclol 麦角生物碱是首例被确认的Cyclol[75]。 环状缩肽沙雷氏菌缩肽同样也是由oxacyclol反应得到的[76]。 用化学方法也得到了类似的环状thiacyclols[77]。 在小分子[78] 和 三肽 [79] 中也发现了经典的azacyclols。多肽是自发地由azacyclol逆转反应得到的[80], Cyclol模型的一个关键推测。数以百计的Cyclol分子已被确认,尽管萊納斯·鮑林的计算说明由于不稳定的高能量,这种分子应该是不存在的[59]。
在林奇长时间致力于X射线晶体学数学运算的过程中,林奇用极大的热情回应了这些关于Cyclol模型及其与生物化学关系的发现[81]。 她出版了两本书,在这两本书中她概括地描述了Cyclol理论和小分子多肽[82][83]。
科学方法示意图
蛋白质结构的Cyclol模型是一个经验证伪作为科学方法的一部分的一个例子。最初的假设的提出是为了解释原理不明的实验现象;而这一假设的结果由实验验明真伪。在这种情况下,肽键的Cyclol结构能强于酰胺键便成为了一个关键的假设。这个假设衍生出一个Cyclol-6分子和Cyclol结构的推测,这反过来说明球状蛋白的半规则多面体模型。一个折叠蛋白质的羰基应该是大量的转变成为羟基的推测是能够通过实验来验证的;然而,光谱分析和化学实验表明,这个推测是不正确的。Cyclol模型还推测了折叠蛋白质中氨基酸的高横向密度,但是这与实验结果不相符。因此,Cyclol模型被推翻,大家开始探求蛋白质结构的新假设,比如20世纪40年代和20世纪50年代提出的α螺旋模型。
有时候有人说Cyclol假设不应该被拓展[84][85], 因为该理论存在先天缺陷,例如其空间位阻,对脯氨酸的不适应性,高自由能都不利于Cyclol反应。虽然这种缺陷使Cyclol假设令人难以置信,但是这并不意味着完全不可能。Cyclol模型是第一个定义明确的球状蛋白结构模型,但是其分子间作用力和蛋白质结构太少被人所知以至于没有被推翻。它巧妙的说明了蛋白质的几个基本特性,同时解释了异常的实验现象。虽然这个理论不正确,Cyclol理论的某些部分却最终被证实,比如Cyclol反应和疏水性相互作用在蛋白质折叠中的作用。一个相对有用的是玻尔模型的氢原子,它从成立之初被认为是不可信的,甚至是它的创造者[86], 推导出了量子力学的最终正确的理论。同样的,萊納斯·鮑林 提出了一个明确定义的DNA模型[87], 一个难以置信却又发人深省的模型[88][89]。 Cyclol理论的故事是一个区域科学发展的例子,提出一个明确定义的假设,测试它,当这个理论被证明是错误的时候排除它。
相反,Cyclol模型是对称性和完美一个不正确的科学理论例子,这两种特性可以被看作“明显正确的”科学理论。例如,沃森—克里克DNA双螺旋模型[89] 有时说是“显而易见”因为其合理的氢键和对称性;不过,另外,在不同的条件下,NDA的不对称结构是有利的[90]。 同样的,阿尔伯特·爱因斯坦的广义相对论的完美理论被认为是不需要实验验证的;然而这个理论需要用量子场论来修正[91]。 Cyclol模型的例子说明,所有的科学理论,即使是最完美,最对称的,也必须通过实验来验证,没有理论是明显正确的,先天成立的,只有更合理的。
文献出处
- Tiselius, A. . Annual Review of Biochemistry. 1939, 8: 155–184. doi:10.1146/annurev.bi.08.070139.001103.
- Svedberg, T. . Nature. 1929, 123 (3110): 871. Bibcode:1929Natur.123..871S. doi:10.1038/123871a0.
- Svedberg, T. . Chemical Reviews. 1934, 14: 1–15. doi:10.1021/cr60047a001.
- Bergmann, M; Niemann C. . Journal of Biological Chemistry. 1937, 118: 301–314.
- Svedberg, T. . Transactions of the Faraday Society. 1930, 26: 741–744.
- Fruton, JS. . Ann. N.Y. Acad. Sci. 1979, 325: 1–18. Bibcode:1979NYASA.325....1F. doi:10.1111/j.1749-6632.1979.tb14125.x.
- Hofmeister, F. . Ergebnisse der Physiologie. 1902, 1: 759–802. doi:10.1007/BF02323641.
- Fischer, E. . Chemiker Zeitung. 1902, 26: 939–940.
- Fischer, E. . Berichte der deutschen chemischen Gesellschaft. 1913, 46 (3): 3253–3289. doi:10.1002/cber.191304603109.
- Sørensen, SPL. . Comptes rendus des travaux du Laboratoire Carlsberg. 1930, 18: 1–124.
- Fruton, JS. . New Haven, CT: Yale University Press. 1999. ISBN 0-585-35980-6.
- Abderhalden, E. . Naturwissenschaften. 1924, 12: 716. Bibcode:1924NW.....12..716A. doi:10.1007/BF01504819.
- Abderhalden, E; Komm E. . Zeitschrift für physiologische Chemie. 1924, 139: 181–204.
- Linderstrøm-Lang, K; Hotchkiss RD, Johansen G. . Nature. 1938, 142 (3605): 996. Bibcode:1938Natur.142..996L. doi:10.1038/142996a0.
- Chick, H; Martin CJ. . Journal of Physiology. 1910, 40: 404–430.
Chick, H; Martin CJ. . Journal of Physiology. 1911, 43: 1–27.
Chick, H; Martin CJ. . Journal of Physiology. 1912, 45: 61–69.
Chick, H; Martin CJ. . Journal of Physiology. 1912, 45: 261–295. - Anson, ML; Mirsky AE. . Journal of General Physiology. 1929, 13: 121–132.
- Anson, ML. . Advances in Protein Chemistry. Advances in Protein Chemistry. 1945, 2: 361–386. ISBN 978-0-12-034202-0. doi:10.1016/S0065-3233(08)60629-4.
- Wu, H. . Chinese Journal of Physiology. 1931, 5: 321–344. Preliminary reports were presented before the XIIIth International Congress of Physiology at Boston (19–24 August 1929) and in the October 1929 issue of the American Journal of Physiology.
- Mirsky, AE; Pauling L. . Proceedings of the National Academy of Science USA. 1936, 22 (7): 439–447. Bibcode:1936PNAS...22..439M. doi:10.1073/pnas.22.7.439.
- Neurath, H; Greenstein JP, Putnam FW, and Erickson JO. . Chemical Reviews. 1944, 34 (2): 157–265. doi:10.1021/cr60108a003.
- Putnam, F. . The Proteins (H. Neurath and K. Bailey, eds.). 1953, 1B: 807–892.
- Astbury, WT; Woods HJ. . Nature. 1931, 127 (3209): 663–665. Bibcode:1931Natur.127..663A. doi:10.1038/127663b0.
- Astbury, WT. . Transactions of the Faraday Society. 1933, 29 (140): 193–211. doi:10.1039/tf9332900193.
- Jordan Lloyd, D. . Biological Review. 1932, 7: 254–273.
- Jordan Lloyd, D; Marriott. . Transactions of the Faraday Society. 1933, 29: 1228.
- Astbury, WT. . Journal of the Textile Institute. 1936, 27: 282–?.
- Wrinch, DM. . Nature. 1936, 137 (3462): 411–412. Bibcode:1936Natur.137..411W. doi:10.1038/137411a0.
- Wrinch, DM. . Nature. 1936, 138 (3484): 241–242. Bibcode:1936Natur.138..241W. doi:10.1038/138241a0.
- Frank, FC. . Nature. 1936, 138 (3484): 242. Bibcode:1936Natur.138..242F. doi:10.1038/138242a0.
- Langmuir, I; Wrinch DM. . Nature. 1939, 143 (3611): 49–52. Bibcode:1939Natur.143...49L. doi:10.1038/143049a0.
- Langmuir, I. . Proceedings of the Physical Society of London. 1939, 51 (4): 592–612. Bibcode:1939PPS....51..592L. doi:10.1088/0959-5309/51/4/305.
- Wrinch, DM. . Philosophical Magazine. 1938, 25: 705–739.
- Wrinch, DM. . Nature. 1936, 142 (3588): 260. Bibcode:1938Natur.142..259.. doi:10.1038/142259a0.
- Dow, RB; Matthews JE, Jr. and Thorp WTS. . American Journal of Physiology. 1940, 131: 382–387.
- Kauzmann, W. . Advances in Protein Chemistry. Advances in Protein Chemistry. 1959, 14: 1–63. ISBN 978-0-12-034214-3. PMID 14404936. doi:10.1016/S0065-3233(08)60608-7.
- Wrinch, DM. . Nature. 1936, 138 (3493): 651–652. Bibcode:1936Natur.138..651W. doi:10.1038/138651a0.
- Wrinch, DM; Jordan Lloyd D. . Nature. 1936, 138 (3496): 758–759. Bibcode:1936Natur.138..758W. doi:10.1038/138758a0.
- Wrinch, DM. . Nature. 1937, 139 (3521): 718. Bibcode:1937Natur.139..718W. doi:10.1038/139718a0.
- Astbury, WT; Wrinch DM. . Nature. 1937, 139 (3523): 798. Bibcode:1937Natur.139..798A. doi:10.1038/139798a0.
- Wrinch, D. . Philosophical Magazine: 64–67.
- Wrinch, DM. . Nature. 1937, 139 (3527): 972–973. Bibcode:1937Natur.139..972W. doi:10.1038/139972a0.
- Wrinch, DM. . Science. 1947, 106 (2743): 73–76. Bibcode:1947Sci...106...73W. PMID 17808858. doi:10.1126/science.106.2743.73.
- Wrinch, DM. . Proceedings of the Royal Society. 1937, A160: 59–86.
Wrinch, DM. . Proceedings of the Royal Society. 1937, A161: 505–524.
Wrinch, DM. . Philosophical Magazine. 1938, 26: 313–332. - Wrinch, DM. . Nature. 1939, 144 (3636): 77. Bibcode:1939Natur.144...77W. doi:10.1038/144077a0.
- Wrinch, DM. . Philosophical Magazine. 1937, 24: 940.
- Wrinch, DM. . Nature. 1938, 142 (3587): 217. Bibcode:1938Natur.142..215.. doi:10.1038/142215a0.
- Wrinch, DM. . Science. 1937, 85 (2215): 566–567. Bibcode:1937Sci....85..566W. PMID 17769864. doi:10.1126/science.85.2215.566.
Wrinch, DM. . Transactions of the Faraday Society. 1937, 33: 1368–1380. doi:10.1039/tf9373301368.
Wrinch, DM. . Journal of the American Chemical Society. 1938, 60 (8): 2005–2006. doi:10.1021/ja01275a514.
Wrinch, DM. . Science. 1938, 88 (2276): 148–149. Bibcode:1938Sci....88..148W. PMID 17751525. doi:10.1126/science.88.2276.148-a.
Wrinch, DM; Langmuir I. . Journal of the American Chemical Society. 1938, 60 (9): 2247–2255. doi:10.1021/ja01276a062.
Langmuir, I; Wrinch DM. . Proceedings of the Physical Society of London. 1939, 51 (4): 613–624. Bibcode:1939PPS....51..613L. doi:10.1088/0959-5309/51/4/306. - Bragg, WL. . Nature. 1939, 143 (3611): 73–74. Bibcode:1939Natur.143...73B. doi:10.1038/143073a0.
Bernal, JD. . Nature. 1939, 143 (3611): 74–75. Bibcode:1939Natur.143...74B. doi:10.1038/143074a0.
Robertson, JM. . Nature. 1939, 143 (3611): 75–76. Bibcode:1939Natur.143...75R. doi:10.1038/143075a0. - Riley, DP; Fankuchen I. . Nature. 1939, 143 (3624): 648–649. Bibcode:1939Natur.143..648R. doi:10.1038/143648a0.
Wrinch, DM. . Nature. 1940, 145 (3687): 1018. Bibcode:1940Natur.145.1018W. doi:10.1038/1451018a0.
Riley, D. . Nature. 1940, 146 (3694): 231. Bibcode:1940Natur.146..231R. doi:10.1038/146231a0. - Neurath, H; Bull HB. . Chemical Reviews. 1938, 23 (3): 391–435. doi:10.1021/cr60076a001.
- Huggins, M. . Journal of the American Chemical Society. 1939, 61 (3): 755. doi:10.1021/ja01872a512.
- Haurowitz, F. . Zeitschrift der physiologischen Chemie. 1938, 256: 28–32.
- Meyer, KH; Hohenemser W. . Nature. 1938, 141 (3582): 1138–1139. Bibcode:1938Natur.141.1138M. doi:10.1038/1411138b0.
- Bergmann, M; Niemann C. . Annual Reviews in Biochemistry. 1938, 7: 99–124. doi:10.1146/annurev.bi.07.070138.000531.
- Neuberger, A. . Proceedings of the Royal Society. 1939, 170: 64–65.
- Neuberger, A. . Nature. 1939, 143 (3620): 473. Bibcode:1939Natur.143..473N. doi:10.1038/143473a0.
- Haurowitz, F; Astrup T. . Nature. 1939, 143 (3612): 118–119. Bibcode:1939Natur.143..118H. doi:10.1038/143118b0.
- Klotz, IM; Griswold P. . Science. 1949, 109 (2830): 309–310. Bibcode:1949Sci...109..309K. PMID 17782718. doi:10.1126/science.109.2830.309.
- Pauling, L; Niemann C. . Journal of the American Chemical Society. 1939, 61 (7): 1860–1867. doi:10.1021/ja01876a065.
- Hotchkiss, RD. . Journal of Biological Chemistry. 1939, 131: 387–395.
- Wrinch, DM. . Journal of the American Chemical Society. 1941, 63 (2): 330–33. doi:10.1021/ja01847a004.
- Wrinch, DM. . Nature. 1940, 145 (3678): 669–670. Bibcode:1940Natur.145..669W. doi:10.1038/145669a0.
- Wrinch, DM. . Science. 1948, 107 (2783): 445–446. PMID 17844448. doi:10.1126/science.107.2783.445-a.
- Wrinch, DM. . Science. 1948, 115 (2987): 356–357. Bibcode:1952Sci...115..356W. PMID 17748855. doi:10.1126/science.115.2987.356.
- Wrinch, DM. . Science. 1948, 116 (3021): 562–564. Bibcode:1952Sci...116..562W. doi:10.1126/science.116.3021.562.
- Wrinch, DM. . Nature. 1939, 143 (3620): 482–483. Bibcode:1939Natur.143..482W. doi:10.1038/143482a0.
- Wrinch, DM. . Nature. 1939, 143 (3627): 763–764. Bibcode:1939Natur.143..763W. doi:10.1038/143763a0.
- Wrinch, DM. . Nature. 1939, 150 (3800): 270–271. Bibcode:1942Natur.150..270W. doi:10.1038/150270a0.
- Anslow, GA. . Physical Review. 1942, 61 (7–8): 547. Bibcode:1942PhRv...61..541.. doi:10.1103/PhysRev.61.541.
- Anslow, GA. . Journal of Applied Physics. 1945, 16: 41–49. Bibcode:1945JAP....16...41A. doi:10.1063/1.1707499.
- Anslow, GA. . Journal of Chemical Physics. 1953, 21 (11): 2083–2084. Bibcode:1953JChPh..21.2083A. doi:10.1063/1.1698765.
- Guedez, T; Núñez A, Tineo E, Núñez O. . Journal of the Chemical Society, Perkin Transactions 2. 2002, 2002 (12): 2078–2082. doi:10.1039/b207233e.
- Bernal, JD. . Nature. 1939, 143 (3625): 663–667. Bibcode:1939Natur.143..663B. doi:10.1038/143663a0.
- Wieland T and Bodanszky M, The World of Peptides, Springer Verlag, pp.193–198. ISBN 0-387-52830-X
- Hofmann, A; Ott H, Griot R, Stadler PA and Frey AJ. . Helvetica Chimica Acta. 1963, 46: 2306–2336.
- Shemyakin, MM; Antonov VK, and Shkrob AM. . Peptides, Proc. 6th Europ. Pept. Symp., Athens. 1963: 319–328.
- Zanotti, G; Pinnen F, Lucente G, Cerrini S, Fedeli W and Mazza F. . J. Chem. Soc. Perkin Trans. 1984, 1: 1153–1157. doi:10.1039/p19840001153.
- Griot, RG; Frey AJ. . Tetrahedron. 1963, 19 (11): 1661–1673. doi:10.1016/S0040-4020(01)99239-7.
- Lucente, G; Romeo A. . Chem. Commun. 1971, ?: 1605–1607.
Rothe M, Schindler W, Pudill R, Kostrzewa U, Theyson R, and Steinberger R. (1971) "Zum Problem der Cycloltripeptidsynthese", Peptides, Proc. 11th Europ. Pept. Symp., Wien, 388–399.
Rothe M and Roser KL. (1988) "Conformational flexibility of cyclic tripeptides", Abstr. 20th Europ. Pept. Symp. Tübingen, p. 36. - Wieland T and Mohr H. (1956) "Diacylamide als energiereiche Verbindungen. Diglycylimid", Liebigs Ann. Chem., 599, 222–232.
Wieland T and Urbach H. (1958) "Weitere Di-Aminoacylimide und ihre intramolekulare Umlagerung", Liebigs Ann. Chem., 613, 84–95.
Brenner, M. Wolstenholme GEW and O'Connor CM, eds., Churchill , 编. . Ciba Foundation Symposium on Amino acids and peptides with antimetabolic activity. 1958. - Wrinch, DM. . Nature. 1957, 179 (4558): 536–537. Bibcode:1957Natur.179..536W. doi:10.1038/179536a0.
Wrinch, DM. . Nature. 1957, 180 (4584): 502–503. Bibcode:1957Natur.180..502W. doi:10.1038/180502b0.
Wrinch, DM. . Nature. 1962, 193 (4812): 245–247. Bibcode:1962Natur.193..245W. PMID 14008494. doi:10.1038/193245a0.
Wrinch, DM. . Nature. 1963, 199 (4893): 564–566. Bibcode:1963Natur.199..564W. doi:10.1038/199564a0.
Wrinch, DM. . Nature. 1965, 206 (4983): 459–461. Bibcode:1965Natur.206..459W. PMID 5319104. doi:10.1038/206459a0. - Wrinch, DM. . Copenhagen: Munksgaard. 1960.
- Wrinch, DM. . New York: Plenum Press. 1965.
- Kauzmann, W. . Protein Science. 1993, 2 (4): 671–691. PMC 2142355
 . PMID 8518739. doi:10.1002/pro.5560020418.
. PMID 8518739. doi:10.1002/pro.5560020418. - Tanford, C; Reynolds J. . Oxford: Oxford University Press. 2001. ISBN 0-19-850466-7.
- Pais, A. . Oxford University Press. 1986. ISBN 0-19-851971-0.
Bohr, N. . Philosophical Magazine. 1913, 26: 1–25. (原始内容存档于2007-07-04).
Bohr, N. . Philosophical Magazine. 1913, 26 (153): 476–502. doi:10.1080/14786441308634993.
Bohr, N. . Philosophical Magazine. 1913, 26: 857–875.
Bohr, N. . Nature. 1914, 92 (2295): 231–232. Bibcode:1913Natur..92..231B. doi:10.1038/092231d0. - Pauling, L; Corey RB. . Proceedings of the National Academy of Sciences. 1953, 39 (2): 84–97. Bibcode:1953PNAS...39...84P. PMC 1063734 . PMID 16578429. doi:10.1073/pnas.39.2.84.
- Franklin, RE; Gosling R. . Nature. 1953, 171 (4356): 740–741. Bibcode:1953Natur.171..740F. PMID 13054694. doi:10.1038/171740a0.
- Watson, JD; Crick F. . Nature. 1953, 171 (4356): 737–738. Bibcode:1953Natur.171..737W. PMID 13054692. doi:10.1038/171737a0.
- Saenger, W. . Springer Verlag. 1988. ISBN 0-387-90762-9.
- Pais, A. . Oxford University Press. 1982. ISBN 0-19-853907-X.
扩展阅读
- "Protein Units Put in Graphic 'Cage'," New York Times, 19 April 1940, page 14.
- "Waffle-Iron Theory of Proteins," New York Times, 2 February 1947, page E9.
- Structures of Matter and Patterns in Science, inspired by the work and life of Dorothy Wrinch, 1894–1976, Proceedings of a Symposium held at Smith College, Northampton, Massachusetts on 28–30 September 1977.Marjorie Senechal, editor. Schenkman Publishing Company, 1980.
- Selected papers of Dorothy Wrinch, from the Sophia Smith Collection (页面存档备份,存于).
- "I Died For Beauty: Dorothy Wrinch and the Cultures of Science," by Marjorie Senechal, Oxford University Press, 2013.