籃尾龍屬
籃尾龍(屬名:,發音:/ˌtæləˈrʊərəs//TAL-ə-ROOR-əs,意為「柳籃般的尾巴」,有時被音譯為塔拉甲龍)是一屬甲龍科恐龍,生存於晚白堊世(約9600萬至8900萬年前)的亞洲。首具化石發現於1948年,之後於1952年由蘇聯古生物學家葉甫根尼·馬列夫命名、敘述,模式種為皺棘籃尾龍(Talarurus plicatospineus)。對於籃尾龍的瞭解來自多具零散的標本,使牠與繪龍一同成為所知最詳盡的甲龍科之一。標本由部分身體骨骼組成,並有五個頭骨被發現及歸入本屬,其中前兩個非常破碎。
| 篮尾龙属 化石时期:晚白堊世 ~ | |
|---|---|
 | |
| 捷克博物館展出的籃尾龍的骨架模型 | |
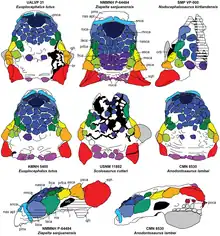
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | †鸟臀目 Ornithischia |
| 科: | †甲龙科 Ankylosauridae |
| 属: | †篮尾龙属 Talarurus Maleev, 1952 |
| 模式種 | |
| †皺棘籃尾龍 Talarurus plicatospineus Maleev, 1952 | |
籃尾龍是種中等體型、結構厚實、陸生、四足的植食性動物,可以生長到5公尺長,體重約介於454至907公斤,將近一噸。如同其他甲龍科,籃尾龍有著厚重的裝甲,尾巴有尾槌,使牠行動緩慢。籃尾龍被分類為甲龍亞目甲龍科的甲龍亞科,屬於一群進階型的甲龍科物種。籃尾龍的化石來自巴彥思楞組,並與同棲地的白山龍有著棲位分化的生活模式,依照牠們嘴部結構的不同,籃尾龍的嘴部為三角形,適合用來刮植。這兩種代表了亞洲已知最古老的甲龍科,但彼此並沒有接近親緣關係。與籃尾龍最近緣的是北美洲的結節頭龍,兩者有著相似的臉部皮內成骨。
許多過去架設的籃尾龍骨架都因為不同因素而過時了,例如:肋骨是朝向身體底部,而非連在身體側面,如同多數甲龍科的情況;不準確的頭骨模型,主要是根據相關物種來重建,而不是拿已知的頭骨材料來當參考;非常突出的四肢;腳部有四趾,但由其他相關甲龍科所示,實際上只有三趾。以上這些錯誤都源於解剖學上的誤解,因為當時對甲龍科還沒瞭解得很全面,而且籃尾龍也沒有完整的骨骼。
發現歷史
籃尾龍的遺骸發現於現在蒙古的戈壁沙漠。正模標本PIN 557-91是在1948年蘇聯-蒙古聯合考察活動期間所發現,從巴彥思楞組貝音詩(Baynshire)化石點的砂質、紅鈣黏土岩之中挖掘出土。籃尾龍於1952年由蘇聯古生物學家葉甫根尼·馬列夫所敘述、命名。PIN 557標本是馬列夫原本指定的正模標本,是一個破碎的頭骨與頭蓋骨後部,包括枕骨區和顱基部、數個脊椎和肋骨、一個肩胛鳥喙骨、一個肱骨、一個尺骨、一個橈骨、一個幾乎完整的手部、一個部分髂骨、一個坐骨、一個股骨、一個脛骨、一個腓骨、一個幾乎完整的腳部、以及關聯的裝甲與鱗甲。屬名Talarurus源自希臘語的τάλαρος("tálaros",意為「籃子」或「柳籃」)和οὐρά ("ourā́",意為「後方」或「尾巴」),意指類似柳籃狀的尾巴末端槌,以及尾巴長度由交錯的骨桿構成,令人聯想到製作柳條籃時的編織方式。種名plicatospineus源自拉丁語的plicātus(意為「摺疊的」)和spīneus(意為「多棘的」或「多刺的」),象徵生前構成裝甲的大量皮內成骨。[1]事實上,這些骨骸是由發現地的六隻個體碎片組合而成。[2]1977年,泰瑞莎·馬瑞沿斯卡考量到使用混雜個體標本會違反規範,於是從中選出PIN 557-91,一個頭骨後部,作為模式標本。與此同時,她將徐龍的種鋸牙徐龍(Syrmosaurus disparoserratus)重新命名為籃尾龍的第二個種鋸牙籃尾龍(Talarurus disparoserratus)。[3]但這個種在1987年被命名為獨立的屬馬列夫龍。[2]所有這些標本都在莫斯科古生物博物館組合成一個骨架模型來展示,雖然非常完整,但在許多方面安裝得非常不準確。[4][5]時至今日,唯一有效的物種只剩下模式種皺棘籃尾龍。[5][6]

現在已知有至少一打的籃尾龍個體標本來自不同的化石點。PIN 3780/1標本於1975年由蘇蒙聯合考察隊採集自巴彥思楞組的貝辛察夫(Bayshin-Tsav)化石點的陸相沉積層,現保存於莫斯科俄羅斯科學院的古生物機構。該標本被歸入籃尾龍,年代為白堊紀土侖階,由頭頂部和破碎的骨骼組成。[7]自2006年以來,在韓國-蒙古聯合國際恐龍計畫的背景下,又有數個額外標本被歸入籃尾龍,發現於貝辛詩和閃俄庫度(Shine Us Khuduk)化石點。這些標本直至2014年都還沒被敘述。[4]另一個來自貝辛察夫化石點、被歸入本屬的標本包含一個尚未敘述的不完整頭骨、頭蓋骨、枕部及腦殼。還有一個採集自巴加塔雅克(Baga Tarjach)化石點的未敘述標本,是個上頜骨碎片與八顆牙齒。Arbour(2014)在研究中列出了許多歸入籃尾龍的額外標本。[4]
後續還有更多標本被發現。在2007年韓蒙國際恐龍考察期間,MPC-D 100/1354、MPC-D 100/1355及MPC-D 100/1356標本採集於戈壁沙漠東部巴彥思楞組的貝音詩化石點。[8]這些標本在2019年被詳細描述,內容包括三個保存程度適中的頭骨,以及關於籃尾龍額外的解剖學及生態學資訊。這些新材料現在永久保存於蒙古科學院的古生物與地質學機構。[6]
敘述
錯誤鑑定
起初馬列夫將籃尾龍描述為腳部有四根腳趾。[1]然而腳部發現的時候關節已脫落;骨架中的重建腳部是用組合而成的,而三根腳趾是比較可能的數字,因為所有其他已知的甲龍類都是三根;早期指稱繪龍也有四根腳趾是錯誤的。另一項假設的特徵也是建立在誤解的基礎上:皮內成骨有皺褶的紋理,構成特別堅固的裝甲,每片鱗甲上都飾有額外的褶皺刺。這些其實是用來保護頸部的半環構造,呈現典型低矮脊狀結構。骨架進一步顯示籃尾龍有著如河馬般的身形,桶狀身軀不像其他甲龍類典型的低矮與寬闊,前肢還強烈外八。這是因為肋骨位置錯誤所造成,使肋骨像是附加上去而不是側向伸出;這個錯誤也造成寬闊的上骨盆無法裝架。[4][5]
體型和鑑定特徵

籃尾龍是中等體型的甲龍類。湯馬斯·霍茨和葛瑞格利·保羅估計身長5公尺;保羅估計體重為2噸,[9]但霍茨估計體重約454至907公斤。[10]如同其他甲龍科,籃尾龍擁有寬闊的嘴喙、沿著身體生長著強韌的皮內成骨,構成大規模的裝甲、還有著名的尾槌。四肢短而強壯,支撐著寬闊、圓潤的腹部。[1][6]Maleev(1956)和Tumanova(1987)提出的鑑定特徵用途受到限制,因為他們主要是在列出籃尾龍與其他甲龍科共有的特徵。維多莉亞·阿布爾建立了單一項自衍徵:頭蓋骨中部的額骨有個隆起的V形區塊;並確認籃尾龍與其他所有甲龍科(結節頭龍除外)之間的差異在於:額骨和鼻骨有著圓錐形、基部為圓形的頭甲(caputegulae)。她還指出腳部實際上有三趾。[4]但Parks等人(2019)提出不同的看法,籃尾龍可透過多項鼻尖上的特徵來鑑定:例如明顯的鼻內頭甲、加長的頭甲、環繞鼻子區域的數個頭甲、圓錐形/多邊形的頭甲…等。[6]
頭骨
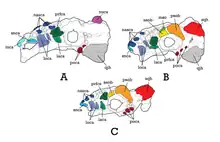
籃尾龍頭骨全長約30公分。[6]正模標本PIN 557-91僅有一個後部頭蓋骨,[3][6]上面有著結節狀頭甲(臉部皮內成骨或甲片),形態類似結節頭龍。Arbour和Currie指出,PIN 3780/1的頭蓋骨頭甲似乎只有細微的不同,不清楚是否代表著物種上的特徵。[5]無論如何,所有能歸入籃尾龍的頭骨都有共同的頭甲形狀(雖然也觀察到一些個體間的細微差異)。[6]最完整的頭骨標本是MPC-D 100/1354,上頜骨前緣保存良好,形狀寬闊、半矩形、癒合、此區域沒有牙齒;可能適用於平坦的表面。頭甲多邊形至結節狀,如同預期假設的樣子,鼻部分布一些頭甲。每個上頜骨有23個齒槽,但牙齒沒有保存下來。頭頂多數頭甲為六邊形,在PIN 557-91的一些是三角形。眼眶由一些長在後部的皮內成骨大幅保護著。MPC-D 100/1355和MPC-D 100/1356的鱗狀骨和方軛骨角厚實且呈三角形,而鱗狀骨更為圓潤。通常籃尾龍的下頜都沒保存下來,MPC-D 100/1355有發現一個關連的右齒骨,但非常破碎。[6]
顱後骨骼

已知大部分的顱後遺骸來自破碎的個體。根據PIN 557,肋骨強烈彎曲且加厚,長度介於50到100公分。脊椎區域有部分遺失,保存下來的有一些頸椎、背椎、尾椎和薦骨。頸椎兩側凹入,椎體加厚,神經弓略高。背椎椎體高且寬,也呈雙凹,帶有特別加長且結構結實的神經弓;有些已斷裂。薦骨比前面部分完整,由4個前薦椎、4個薦椎以及第一節尾椎組成,這些脊椎癒合在一起,附著一些肋骨,尺寸大致由後向前增加。有些單獨的尾椎存在,似乎表明尾部由25至30節尾椎構成。最前面的尾椎具有某種程度上加長的神經弓和椎體,人字骨癒合呈三角形。相較之下,最後面的尾椎有非常扁平的人字骨和神經弓。[1][11]因為標本僅部分保存下來,實際上的皮內成骨排列方式是未知的,然而還是有發現一些。保存的皮內成骨非常零散,包括一些頸部半環和棘狀身體皮內成骨。皮內成骨非常結實,多數棘狀身體皮內成骨測量高15公分。[1][11]MPC-D 100/1355被發現與一塊部分半環與其他身體部位相連。[6]
手部只有左邊保存下來,幾乎完整,保存了五個手趾,只缺少一些爪和指骨。保存的左邊腳部非常獨特,被敘述成有五根腳趾,[1][11]但這後來被證明是最初架設的骨架所造成的推論,三根腳趾才是比較正確及可能的,如同其他相關甲龍科所見。[5]整體來說,手部和腳部的蹠骨都非常厚實。胸帶由一條長約60公分的肩胛鳥喙骨、以及肱骨、尺骨和橈骨組成,這些部位與顱後其他部分比起來非常結實,屬於左臂。左肩胛鳥喙骨非常結實,且帶有一個發育良好給肱骨附著的肩臼,雖然肩胛骨某種程度上長度縮短。肱骨末端非常寬闊與加厚,長33.5公分;與甲龍的有些相似之處。骨盆帶很破碎,包括部分壓扁的髂骨和坐骨。後肢材料包括右股骨與左脛骨。保存的股骨直且非常結實,大轉子與小轉子癒合;測量長47公分。脛骨較短但非常寬,全長29.8公分。此外有一個腓骨也保存下來,長26.7公分;腓骨整體上比脛骨和股骨還直和細,遠端寬於近端。[1][11]
分類
甲龍科的系統發生學關係難以確認,因為對許多物種的瞭解僅來自部分骨骼,實際的裝甲位置很少能保存下來,癒合的皮內成骨使得許多頭骨細節變得模糊不清,而且甲龍科的顱後骨骼變化不大,脊椎、骨盆、四肢只有很細微的差異。籃尾龍過去被認為是已知年代最古老的甲龍科,並有著一些與結節龍科共同的基礎特徵,這些特徵在進階型甲龍科消失,例如四根腳趾。然而,這些「假設的」原始特徵被證實是最初重建骨骼時的人工產物。近期的系統發生學分析提供了一些將籃尾龍分類為甲龍亞科(進階型甲龍科)的證據。可以藉此推論整個甲龍科演化支早在侏儸紀時期就已經出現,此理論將與籃尾龍相對較古老的年代相呼應;Coombs(1978)曾提出甲龍科與結節龍科互成姊妹群(在所有多刺甲龍亞科都屬於結節龍科的前提下),可能可以用以上理論來支持。[12][5]
在最初的敘述論文(Maleev, 1952)中,將籃尾龍分類為徐龍科(Syrmosauridae)。[1]Walter Preston Coombs(1978)認為籃尾龍與包頭龍是相同的物種,但後來的研究都不支持此理論。[12]Maryańska(1977)指出籃尾龍與包頭龍的差異,包含:頭骨形狀、甲板形態、四根腳趾。[3]Vickaryous等人(2004)提到白堊紀晚期存在兩個不同的甲龍科演化支,一個是北美洲物種組成,另一個僅包含亞洲物種。[13]但在Arbour(2014)製作的演化樹中,籃尾龍與北美洲甲龍科較近緣,而相對地離其他亞洲甲龍科較遠。其中一些呈現出籃尾龍與結節頭龍互成姊妹群。[4]
| 甲龍亞科 Ankylosaurinae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
古生物學
2007年的頭骨標本MPC-D 100/1354,是個保存良好、幾乎完整的腦殼,為籃尾龍的顱內神經構造和飲食習性提供了新的資訊。Paulina-Carabajal等人(2017)將MPC-D 100/1354與一個非常完整的多智龍頭骨進行全面性細節的描述,根據對所選標本顱內膜區域的檢驗,他們指出甲龍科有著發育良好的視覺穩定性以及聽覺感官,這與結節龍科相當不同。最早由Maryańska(1977)[3]提到小腦鬈存在於籃尾龍,後來也在其他有關的甲龍科發現,但這個小葉構造在結節龍科似乎已經退化甚至消失。在多數甲龍科中,小腦鬈都相對很大。另一項神經解剖學特徵是加長的聽壺,這在包頭龍、多智龍和籃尾龍都非常明顯。這些解剖學特徵顯示甲龍科擁有很大範圍的聲音感知度,特別是低頻震動。[8]除此之外,MPC-D 100/1354保存的嘴喙寬闊,形狀幾乎呈矩形且大致厚實。籃尾龍厚實的嘴喙可能可以有效率的處理平原地低矮處的植物。證據顯示籃尾龍有著刮植者的覓食習性。[6]
古生態學

籃尾龍的化石遺骸出土於巴彥思楞組的沉積層,年代被認為可追溯至晚白堊世森諾曼階至三冬階,約9600萬至8900萬年前。[14]根據Park等人(2019),籃尾龍佔據刮植者的植食性動物生態棲位,而同一地層的另一種甲龍科白山龍則填補攝植者的棲位。舉例來說,籃尾龍有著寬闊矩形的嘴喙,適合吃低矮的植物;白山龍的嘴喙加長鏟狀,適合吃高處植物。為了說明呈現這樣的差異性覓食策略,研究人員將甲龍科與現生的黑犀牛、白犀牛進行比較;白犀牛比較像籃尾龍,吻部寬闊矩形,特化來吃草;另一方面,黑犀牛的嘴巴尖,像白山龍一樣適合攝植。有了這些解剖學特徵和生態學實例,很明顯可以證實這些甲龍科有著棲位分化。[6]巴彥思楞組還可以觀察到另一個棲位分化的例子是鐮刀龍科的死神龍和慢龍。[15]

晚白堊世的化石點,如貝音詩和貝辛察夫[16],與籃尾龍共享棲地的巴彥思楞組上層動物群包含高度多樣性的恐龍種群,如大型馳龍科阿基里斯龍[17];鐮刀龍科的死神龍[18]、慢龍[18];似鳥龍類的似金翅鳥龍[19];小型可能未成年的角龍類雅角龍[20];小型鴨嘴龍類戈壁鴨龍[16];以及對立生態棲位的白山龍[21]。根據鈣質層、河流、湖泊沉積物判斷,巴彥思楞組是個大型陸相半乾旱氣候地區,充斥著曲流和湖泊群,[22][23]全區域大量分布著被子植物和裸子植物。[24][25]依照一些亞洲地層的生物地層學模式,巴彥思楞組可能與二連組有關聯性。[26][27]
參考來源
- Maleev, E. A. [A new ankylosaur from the Upper Cretaceous of Mongolia] (PDF). Doklady Akademii Nauk SSSR. 1952, 87 (2): 273–276 [2021-03-14]. (原始内容存档 (PDF)于2020-11-05) (俄语).
- Tumanova, T. A. (PDF). The Joint Soviet-Mongolian Paleontological Expedition Transaction. 1987, 32: 1–76 [2021-03-14]. (原始内容存档 (PDF)于2021-01-10) (俄语).
- Maryańska, T. (PDF). Palaeontologia Polonica. 1977, 37: 85–151 [2021-03-14]. (原始内容 (PDF)存档于2020-07-12).
- Arbour, V. M. (PDF). Ph.D thesis. 2014: 265 [2021-03-14]. (原始内容存档 (PDF)于2021-04-13).
- Arbour, V. M.; Currie, P. J. . Journal of Systematic Palaeontology. 2015, 14 (5): 385–444. doi:10.1080/14772019.2015.1059985.
- Park, J. Y.; Lee, Y. N.; Currie, P. J.; Kobayashi, Y.; Koppelhus, E.; Barsbold, R.; Mateus, O.; Lee, S.; Kim, S. H. . Cretaceous Research. 2019. doi:10.1016/j.cretres.2019.104340.
- Kurzanov, S. M.; Tumanova, T. A. . Paleontologicheskii Zhurnal. 1978, 3: 369–374 (俄语).
- Paulina-Carabajal, A.; Lee, Y. N.; Kobayashi, Y.; Lee, H. J.; Currie, P. J. . Palaeogeography, Palaeoclimatology, Palaeoecology. 2017, 494. Bibcode:2018PPP...494..135P. doi:10.1016/j.palaeo.2017.11.030.
- Paul, G. S. . Princeton, New Jersey: Princeton University Press. 2010: 231. ISBN 978-0-6911-3720-9.
- Holtz, T. R.; Rey, L. V. . Random House. 2007. Genus List for Holtz 2012 (页面存档备份,存于) Weight Information (页面存档备份,存于)
- Maleev, E. A. [Armored Dinosaurs of the Upper Cretaceous of Mongolia Family Ankylosauridae] (PDF). Trudy Paleontologicheskogo Instituta Akademiy Nauk SSSR. 1956, 62: 51–91 [2021-03-21]. (原始内容存档 (PDF)于2020-11-05) (俄语).
- Coombs, W. P. (PDF). Palaeontology. 1978, 21 (1): 143–170 [2021-03-16]. (原始内容存档 (PDF)于2022-08-23).
- Vickaryous, M. K.; Maryańska, T.; Weishampel, D. B. . . University of California Press. 2004: 363–392. ISBN 978-0-5202-5408-4.
- Kurumada, Y.; Aoki, S.; Aoki, K.; Kato, D.; Saneyoshi, M.; Tsogtbaatar, K.; Windley, B. F.; Ishigaki, S. . Terra Nova. 2020. doi:10.1111/ter.12456.
- Zanno, L. E.; Tsogtbaatar, K.; Chinzorig, T.; Gates, T. A. . PeerJ. 2016, 4: e1885. PMC 4824891
 . PMID 27069815. doi:10.7717/peerj.1885.
. PMID 27069815. doi:10.7717/peerj.1885. - Khishigjav Tsogtbaatar; David B. Weishampel; David C. Evans; Mahito Watabe. . PLoS ONE. 2019, 14 (4): e0208480. Bibcode:2019PLoSO..1408480T. PMID 30995236. doi:10.1371/journal.pone.0208480.
- Perle, A.; Norell, M. A.; Clark, J. . Contributions from the Geology and Mineralogy Chair, National Museum of Mongolia. 1999, (101): 1–105. OCLC 69865262.
- Barsbold, R.; Perle, A. (PDF). Acta Palaeontologica Polonica. 1980, 25 (2): 190–192 [2021-03-16]. (原始内容存档 (PDF)于2017-03-12).
- Kobayashi, Y.; Barsbold, R. (PDF). Canadian Journal of Earth Sciences. 2005, 42 (9): 1501−1521 [2021-03-16]. Bibcode:2005CaJES..42.1501K. doi:10.1139/e05-044. hdl:2115/14579. (原始内容存档 (PDF)于2015-10-01).
- Sereno, P. C. . (PDF). Cambridge University Press. 2000: 489–491 [2021-03-16]. (原始内容存档 (PDF)于2019-12-25).
- Tumanova, T. A. [A new armored dinosaur from Southeastern Gobi]. Paleontologicheskii Zhurnal. 1993, 27 (2): 92–98 (俄语).
- Jerzykiewicz, T.; Russell, D. A. . Cretaceous Research. 1991, 12 (4): 345–377. ISSN 0195-6671. doi:10.1016/0195-6671(91)90015-5.
- Hicks, J. F.; Brinkman, D. L.; Nichols, D. J.; Watabe, M. . Cretaceous Research. 1999, 20 (6): 829–850 [2021-03-16]. doi:10.1006/cres.1999.0188. (原始内容存档于2020-10-18).
- Khand, Y.; Badamgarav, D.; Ariunchimeg, Y.; Barsbold, R. . . 2000: 49–79. doi:10.1016/s0920-5446(00)80024-2.
- Ksepka, D. T.; Norell, M. A. (PDF). American Museum Novitates. 2006, (3508): 1–16 [2021-03-16]. (原始内容存档 (PDF)于2021-08-31).
- Averianov, A.; Sues, H. (PDF). Journal of Stratigraphy. 2012, 36 (2): 462–485 [2021-03-16]. (原始内容 (PDF)存档于2019-03-07).
- Tsuihiji, T.; Watabe, M.; Barsbold, R.; Tsogtbaatar, K. . Cretaceous Research. 2015, 56: 60–65. doi:10.1016/j.cretres.2015.03.007.