细胞迁移
细胞迁移,与细胞移动同义,与细胞运动義近,指的是细胞在接收到迁移信号或感受到某些物质的浓度梯度后而产生的移动。移動过程中,细胞不断重复着向前方伸出突觸/偽足,然后牵拉後方胞体的循环过程。细胞骨架和其结合蛋白,還有細胞間質是这個过程的物质基础,另外还有多种物质會对之进行精密调节。

若以移動方式與型態來比較,细胞迁移是通过胞体形变进行的定向移动,这有别于其他;如细胞靠鞭毛与纤毛的运动、或是细胞随血流而发生的位置变化,而且就移動速度來看,相比起后两者,细胞迁移要慢得多。舉例而言:成纤维细胞[註 1]的移动速度为1微米每分,若以精子的平均游動速度56.44微米/每秒,即3384微米/每分[1]來比較,兩者差距約3000倍以上。角膜细胞即使比成纤维细胞快上十倍,但是要完成从不来梅到汉堡这93公的路程仍需要17123年[註 2]。而且细胞用力甚轻。成纤维细胞胞体收缩的力只有2×10−7牛顿,而角膜细胞的则是2×10−8牛顿(一牛顿约为人用手举起一鸡蛋所用的力道)[2]。
但此等「步缓力微」的細胞遷移,却是细胞觅食、傷口痊癒、胚胎發生、免疫反應、感染和癌症转移等等生理现象所涉及到的。因此细胞迁移是目前细胞生物学研究的一个主要課題,科学家們试图通过对细胞迁移的研究,在阻止癌症转移、異體植皮等医学应用方面取得更大成果。也因為細胞遷移獨有的运动特性,成為今生物学熱門研究方向。
细胞迁移的研究史
1675年,显微技术的先驱人物安东尼·凡·列文虎克往英国皇家学会寄出一封信,里面描写了细菌的运动。这封信可以说是打开了科学家对细胞迁移研究的第一页。在往后这300多年时间,人们就一直试图去理解细胞迁移过程的细节。
而细胞迁移的关键物质—细胞骨架则要等到20世纪才被发现。虽然1939年科学家阿尔伯特·山特吉尔吉就已发现细胞骨架的成分—肌动蛋白和肌球蛋白,但是因为电子显微镜制作样本时需要对样品进行0到4 °C的低温固定,在这样的温度下细胞骨架会被破坏,即所谓的“解聚”。所以当时认为细胞质不过是一“蛋白汤”,各种细胞器悬浮于细胞质液()中。但在60年代后,人们使用戊二醛常温固定的方法开始逐渐发现细胞骨架。科学家发现,细胞骨架在细胞迁移的过程中起到承载和支撑的作用。在20世纪末21世纪初,科学家对细胞迁移复杂机理的认识有了非常大的进步,对细胞与基质的粘着,非对称性极化和胞内分层运动都有了进一步的了解。但是整个过程其实仍未被了解透彻,很多中间过程就是连起作用的物质都未明。科学家对其中部分需要进行假设,再进一步通过实验去证实。
研究技术
为了研究某一蛋白质在细胞迁移中所扮演的角色,一般来说科學家可以将某蛋白的编码基因进行突变,甚至应用新近的RNAi现象,或者加入该蛋白质的阻断剂()来抑制某一個蛋白質的表現,並分析此抑制對於細胞遷移的影響,反而得知被抑制的蛋白質與細胞遷移的作用。
新科技对细胞迁移研究起到了极大的推动作用。科学家通过ECIS技术(;電子細胞基質阻抗判斷)可以观察到细胞在传统细胞培养甚至是液体环境中的移动行为[3]。根据ECIS技术观测细胞电学参数的能力,ECIS技术还可以量化测量肿瘤细胞迁移过程中细胞层形态变化[4]。同样是在肿瘤研究领域,ATIM(Fluorescence- Assisted Transmigration Invasion and Motility Assay,荧光协助转移侵入和运动分析法) 提供了快速定量细胞侵入(細胞從一個區域進入另一區域)的更好方法,允许检测大量样品和不同条件下时间依赖性侵入。更重要的是,这一系统可以方便地通过在多孔膜上增加胞外基质的厚度来监测细胞侵入结构的深度[5]。韩国延世大学的朴宗哲和朴峰珠则发展出一套细胞跟踪系统。它是由计算机辅助的时间流逝显示微观复制系统,其中有影象形成过程软件,其程序编制含有自动影象分析和自设计CO2微小细胞培育器,它的功能是在一个倒置显微镜平台上,对于细胞迁移进行迅速而精确的分析,从而形成對於细胞的培育。目前已知他们运用这一计算机辅助系统计算了外细胞间质(ECMs)覆盖面的细胞迁移过程[6]。
斑马鱼是目前在该领域最常用于研究的生物。细胞迁移是脊椎动物胚胎發育的核心过程之一。细胞从原分裂生成的部位移动到目的部位就是细胞的迁移。斑马鱼有着很大的优势,首先是其胚胎能在母体外发育,速度快,受精24小时后身体的器官已大部分就位。而且斑马鱼繁殖量大,容易对之进行变异。还有其胚胎透明,在高分辨率快进摄影技术的帮助下,人们可以很好的观察到细胞迁移的过程,还可以利用绿色荧光蛋白(GFP)可以观察到细胞在斑马鱼体内的分布情况[7]。
参与细胞迁移的分子
细胞迁移需要内外因素的配合。外部的因素指的是细胞外的信号分子。内部因素则指细胞的信号传导系统和执行运动的细胞骨架和分子马达,还有参与粘着斑形成的各种分子(关于参与形成粘着斑的各种分子请见突出与底质的粘着)。细胞外信号结合胞膜受体完成其使命后,需要细胞内信号分子接力,将运动信息进一步传给细胞迁移的执行单位——细胞骨架和分子马达。种类繁多的细胞内信号分子会相互作用,影响后述这两种分子的分布,结构和活性,达到精细调整细胞运动的目的。
细胞外
在一定条件下,细胞外的化学信号能引发细胞的定向移动。这些信号有些时候是底质表面上一些难溶物质,有些时候则是可溶物质。信号分子有很多,可以是肽,代谢产物,细胞壁或是细胞膜的残片,但是作用方式却是一样的,就是与细胞膜表面上的受体结合,启动细胞内信号,完成一系列的反应,去激活或抑制肌动蛋白结合蛋白的活性,最终改变细胞骨架的状态。可溶物质通常不是均匀溶解在溶剂中,而是靠近源的区域浓度高,远离源的区域浓度低,形成所谓的“浓度梯度”。细胞膜上的受体可感受到那些被称为化学趋向吸引物(),并且逆着它们的浓度梯度去追根寻源。某些信号分子甚至会影响细胞移行的速度,这些信号分子则被称为化学趋向剂()。细胞这种因化学分子改变自己移动的行为,被称为化学趋向性。例如盘基网柄菌(Dictyostelium discoideum)会逆着cAMP浓度梯度的运动。白血球也会受到一些细菌分泌的三肽化学物质f-Met-Leu-Phe(N-甲酰蛋-亮-苯丙氨酸)吸引而往细菌移动,发挥其免疫功能。而在胚胎发生中的神经嵴细胞则并非靠浓度梯度,而是路标物质识别其去向(请见下文“路标信号”一节)。
但是细胞外基质中也存在着一些蛋白,如硫酸软骨蛋白多糖(chondroitin sulfate proteoglycan)会与神经细胞的粘着蛋白起作用,对细胞迁移形成阻滞。它会抑制脊髓损伤患者神经损伤区域新突触的相连与再生[8][9]。
细胞内

细胞外信号种类繁多,但是当它们与细胞膜上受体结合之后,细胞内起作用的途径却只有有限的几种。而与细胞迁移有关的信号传导过程如下:信号分子结合到膜上受体,或者是激活与受体偶联的蛋白质—大G蛋白,或者先是激活受体酪氨酸激酶,再激活下游的小G蛋白Ras。G蛋白是一个很大的家族,包括Rho,Rac,Ras等小家族,它们在细胞中扮演着信号传导开关的角色。当它们与GDP结合时,呈现失活状态。在鸟嘌呤交换因子(英文:Guanin exchange factor,简称GEF)的帮助下,G蛋白脱离GDP并与GTP结合,进入激活状态。G蛋白的GTP会被GTP酶激活蛋白(英文GTPase-activating proteins,简称GAP)水解,并释放出其中的能量,让G蛋白行使其功能。就是说,G蛋白通过这一GTP/GDP循环在激活/失活状态中回旋,传递信号。当G蛋白被激活后,它下游的多种分子会被激活。从插图2中可见,这些下游分子本身会形成网络,相互制约,或者是相辅相承。它们调控着细胞迁移中各个方面。它们作用的详细情况请见文章中的相应章节。
而致癌物质也可以通过这些信号传导通路发挥其负面作用,如强烈致癌物质佛波酯(Phorbol ester)。佛波酯会不可逆地激活细胞的RasGRP3/4,以激活Ras,Ras会再激活蛋白激酶C(Protein kinase C,PKC)。后者是调节细胞分裂和分化的酶。它被佛波酯不正常的激活,有可能对癌症的产生起促进作用。研究还发现,佛波酯对黑素瘤(melanoma)细胞转移到肺部有促进作用[10]。而细菌者,如志贺氏菌会在宿主胞膜上打洞,向细胞质注入效应蛋白质,激活宿主Rac和Cdc42,调整细胞的微丝网络,以使自己顺利进入宿主内。
细胞骨架
细胞骨架的定义分为狭义和广义两种,前者是微丝,微管和中间纤维的总称,它们存在于细胞质内,又被称为“胞质骨架”。后者还包括细胞外基质(extracellular matrix)、核骨架(nucleoskeleton)和核纤层(nuclear lamina)。细胞骨架是细胞内运动,细胞器固定,细胞外型维持,信号传导和细胞分裂的物质基础之一。
微丝和其结合蛋白

微丝是由肌动蛋白(Actin)组成的直径约为7nm的纤维结构。肌动蛋白单体(又被称为G-Actin,全称为球状肌动蛋白,Globular Actin,下文简称G肌动蛋白)为球形,其表面上有一ATP结合位点。肌动蛋白单体一个接一个连成一串肌动蛋白链,两串这样的肌动蛋白链互相缠绕扭曲成一股微丝。这种肌动蛋白多聚体又被称为纤维形肌动蛋白(F-Actin,Fibrous Actin)。
微丝能被组装和去组装。当单体上结合的是ATP时,就会有较高的相互亲和力,单体趋向于聚合成多聚体,就是组装。而当ATP水解成ADP后,单体亲和力就会下降,多聚体趋向解聚,即是去组装。高ATP浓度有利于微丝的组装。所以当将细胞质放入富含ATP的溶液时,细胞质会因为微丝的大量组装迅速凝固成胶。而微丝的两端组装速度并不一样。快的一端(+极)比慢的一端(-极)快上5到10倍。当ATP浓度达一定临界值时,可以观察到+极组装而-极同时去组装的现象,被命为“踏车”。
微丝的组装和去组装受到细胞质内多种蛋白的调节,这些蛋白能结合到微丝上,影响其组装去组装速度,被称之为微丝结合蛋白(association protein)。
微丝的组装先需要“核化”(nucleation),即几个单体首先聚合,其它单体再与之结合成更大的多聚体。Arp复合体(Arp:Actin related-protein)是一种能与肌动蛋白结合的蛋白,它起到模板的作用,促进肌动蛋白的多聚化。Arp复合体由Arp2,Arp3和其它5种蛋白构成,也寫成Arp2/3複合體。
封闭蛋白(end-blocking protein)则是微丝两端的“帽子”。当这种蛋白结合到微丝上时,微丝的组装和去组装就会停止。这对一些长度固定的蛋白来说很重要,如细肌丝。
而前纤维蛋白(Profilin,或译G肌动蛋白结合蛋白)则是促进多聚的,相应地促解聚的蛋白则有丝切蛋白(Cofilin)。纤丝切割蛋白(filament severing protein),如凝溶胶蛋白(Gelsolin),能将微丝从中间切断。粘着斑蛋白(Vinculin)则能固定微丝到细胞膜上,形成粘着斑。交联蛋白(cross-linking protein)有两个以上肌动蛋白结合位点,起到连接微丝的作用,其中,丝束蛋白(fimbrin)帮助微丝结成束状,而细丝蛋白(filamin)则将微丝交联成网状。
微管
微管是另一种具有极性的细胞骨架。它是由13 条原纤维(protofilament)构成的中空管状结构,直径22—25 nm。每一条原纤维由微管蛋白二聚体线性排列而成。微管蛋白二聚体由结构相似的α和β球蛋白构成,两种亚基均可结合GTP,α球蛋白结合的GTP 从不发生水解或交换,是α球蛋白的固有组成部分,β球蛋白结合的GTP 可发生水解,结合的GDP 可交换为GTP,可见β亚基也是一种 G 蛋白。微管和微丝一样,具有生长速度较+端和较慢的-端。微管在细胞内起支撑作用。另外它还是两种运载分子:驱动蛋白(Kinesin)和发动蛋白(Dynein),的行走轨道。微管,可能连带负在其上的发动蛋白会发放信号促进粘着斑的解聚,后者是粘着斑的周转和尾部与底质分离过程中重要的一步。
中间纤维
中间纤维(intermediate filaments,IF)直径10nm 左右,介于微丝和微管之间。与后两者不同的是中间纤维是最稳定的细胞骨架成分,它主要起支撑作用。中间纤维在细胞中围绕着细胞核分布,成束成网,并扩展到细胞质膜,与质膜相连结。中间纤维没有正负极性。
角蛋白是中间纤维中的一类,分子量约40~70KD,出现在表皮细胞中,在人类上皮细胞中有20 多种不同的角蛋白,分为α 和β 两类。角蛋白赋予细胞体一定的刚性。癌细胞需要对角蛋白进行重新分布,以使自身变得柔韧,可以通过基底膜或血管壁上的细小孔洞。
分子马达

分子马达(Motorprotein)是一类蛋白质,它们的构象会随着与ATP和ADP的交替结合而改变, ATP水解的能量转化为机械能,引起马达形变,或者是它和与其结合的分子产生移动。就是说,分子马达本质上是一类ATP酶。例如肌肉中的肌球蛋白(Myosin)会拉动粗肌丝向中板移动,引起肌肉收缩。而另外两种分子马达:驱动蛋白(Kinesin)和动力蛋白(Dynein),它们能够承载着分子“货物”——如质膜微粒,甚至是线粒体和溶酶体,在由微管构成的轨道上滑行,起到运输的作用。例如驱动蛋白的重链则会运输参与粘着斑解聚过程的信号物质。所以在驱动蛋白的重链受到抑制的情况下,粘着斑会比正常情况下显得更大。
肌球蛋白是微丝结合蛋白,最早发现于肌肉组织,1970年代后逐渐发现许多非肌细胞的肌球蛋白。其家族有13个成员,每个成员在结构上都分为头,颈和尾部三个部分,形似豆芽,而组成上则有轻重两种链。其中的调节轻链(regulatory light chain)是肌球蛋白接受调解的位点,就是说,调节轻链的磷酸化/去磷酸化状态影响着肌球蛋白的活性。其中I和II型是研究得最彻底的分子马达。一些细胞具有突变的肌球蛋白,它们能正常伸出伪足,但是却不能成功移动。I型肌球蛋白是单体,II型和V型则是二聚体。趋向微丝的+极运动。蛋白的头部能就尾部作屈伸运动,并在“屈”的时候拉动微丝相对向后运动。肌球蛋白除了参与肌肉收缩外,还被认为是细胞迁移所需的重要分子之一。肌球蛋白非常可能参与了“前进的四个步骤”里面胞体收缩一步。另外,在细胞突出一端也可观察到肌球蛋白,它可能是帮助运输粘着所需要的蛋白质,提高粘着效率。
过程簡述
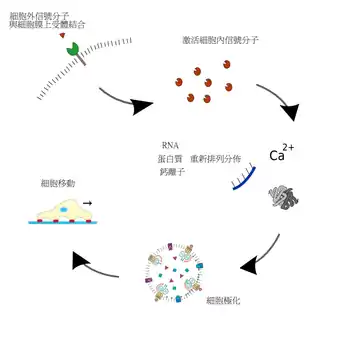
細胞遷移的過程可以用右图阐明。细胞迁移是一系列生理程序的集合,接收到外界信号后(关于外界信号作用于细胞的过程,请见运动方向的确定和极化),细胞会作出相应的层级反应(),其中的每一个阶段都要相应的蛋白质在适当的位置被激活。这一连串的蛋白质的活化并不是同时平行进行,而是有先后顺序的。处于悬浮状态的成纤维细胞,会处于一种所谓闲逛(random walk)状态,或者被称之为处于各向同性伸展期(isotropic spreading phase),它在不断伸出伪足后又不断将之收回,可能是要在就近一探其究竟。细胞或者是靠外界信号物質濃度梯度(请见化学趋向性),或是利用某些特定分子作為路标信号,确定前进的方向。細胞內部的分子会因应需要發生變化,一些蛋白质和离子会重新排列,显示出不均匀分布,就是出现了所谓的极性,而这个过程请见極化。
值得一提的是,细胞在前进的过程中,可以不断改变其前进的方向。在显微镜下观察大肠杆菌(Escherichia coli)寻找食物时的运动,可见细胞先向前直线移动一段时间,然后会停下来并且调整一下方向,然后又再作直线移动。如此不断反复。可见细胞内调控能力的有效和精确。
细胞极化后,細胞的前端會伸出極狀足(请见细胞前端突出)。极状足伸出后,會與細胞前方的底質附著;粘着处会形成一种固定结构,名曰粘着斑(请见突出与底质的粘着)。此时,胞体主体會被牵拉向前(请见细胞体前移);最後細胞的後端与底质剥离(请见牵引尾部往前)。这样前进的4個步驟完成,并准备下一次循环。
不同的细胞,它们直线运动的速度和持续的时间是不同的。使用分子干扰技术可以很好的研究这两者。一般来说,细胞直线前进速度越慢,其保持直线运动的时间就越长,例外是鱼的上皮细胞,它能够在快速迁移的同时,显示出长时间保持直线运动的能力。
過程詳細機制
运动方向的确定和极化
细胞要“有效”的运动,首先要确定哪里是“前方”,否则会出现细胞体两部分同时向相反方向运动的情况(这在一些神经系统不发达的无脊椎动物中是可以见到的)。而这个确定方向的过程,其实是细胞内与运动有关的物质,如钙离子,肌球蛋白,和各类细胞骨架物质的浓度重新分布的过程,因此细胞显出“极性”,这个重新分布的过程也因而被称为细胞“极化”(Polarization)。细胞可以靠其表面的受体顺着诱饵的浓度“嗅”到其发放的源头,或者“感受到”底质上的“路标”而确定自身去向,前者被称为细胞的化学趋向性。这两种机制好比一个人或者是顺着楼梯行进或是依着路标的指示驾驶一样。
化学趋向性
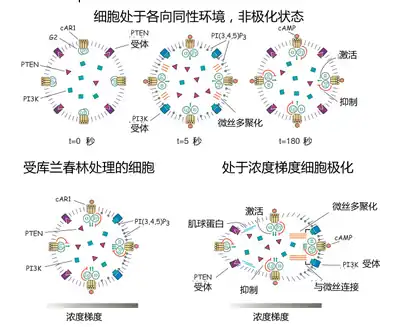
细胞的化学趋向性是指细胞具有沿着外界某一物质的浓度朝向或是背离发放源运动,甚至形变的能力,这是一种基本的细胞反应。例如黏菌盘基网柄菌(Dictyostelium discoideum)会凭这种能力去寻找外界的养料,哺乳类动物细胞在这方面的行为和它很相似。当细胞饥饿时,它会极化并且在周围寻找环磷酸腺苷(3,5-cyclic adenosine monophosphate),简称cAMP。細胞膜上有四种cAMP受体,分别被命名为cAR1—cAR4。它们和膜内其中的“分子开关”——三联体大G蛋白(G Protein,三个亚基被称为α,β和γ)相偶联。这里所说的G蛋白是其4大家族里面的Gi家族。该家族的作用是抑制cAMP的生成。但是也可在其他哺乳类细胞里面看到其它的信号传导策略,例如cGMP,Ca2+和IP3水平的短暂上升。而三磷酸磷脂酰肌醇(Phosphatidylinositol trisphosphate),简称PI(3,4,5)P3则是阿米巴和白细胞极化过程中的枢纽物质。
在细胞接收到外部迁移信号cAMP之前,一部分同源性磷酸酶-张力蛋白(phosphatase and tensin homolog,由第十染色体同源丢失性磷酸酶-张力蛋白基因,即PTEN基因编码,其英文全名为phosphatase and tensin homolog deleted on chromosome ten,又名同源磷酸酶-张力蛋白基因(变异于多种进行性癌1),英文:phosphatase and tensin homolog (mutated in multiple advanced cancers 1)。但下文所提及的PTEN,乃指其翻译产物,即同源性磷酸酶-张力蛋白,而非基因。[11][12])分布在细胞膜上。而磷脂酰肌醇-3激酶(下文将之简称为PI3K,Phosphoinositide 3-Kinase)则分布在细胞质裏。当细胞膜上的受体与外界的化学信号结合后,分子开关G蛋白被激活,α亚基携带着一GTP并与βγ亚基分离。通过这种形式,信号被进一步传递,结果是在胞质内的PI3K迅速结合到细胞膜上,而PTEN则从细胞膜上脱离。这两种分子的位置变化带来的是PI(3,4,5)P3的浓度上升。
PI(3,4,5)P3的浓度会因为细胞内一套负反馈机制被压制下去。而细胞前端比后端接触到更多的外界刺激,因此PI(3,4,5)P3会在集中到前端。PI(3,4,5)P3浓度的上升则会促进所在部位微丝的多聚化,细胞移动方向也就被确定下来了。
路标信号

信号分子可能并不一定要形成浓度梯度才能为细胞指路,或者只要它做出连续的分布让细胞“顺瓜摸藤”即可,甚至是沿途的不迁移细胞,在自身胞膜表面表达一些蛋白质,做出“邀请”或是“排挤”的姿态。迁移中的细胞被观察到会不断伸出伪足“摸索”其周围的环境,找出与其膜上受体配对的信号分子后,经过一番“吸引—排斥”的拉锯拔河,得出净迁移方向后再继续前进。细胞对路径的识别可能涉及多种分子和机制。不同的细胞可能使用不同的分子组合作迁移指引。 德国图宾根马克思·普朗克研究所为了了解细胞究竟是凭哪些信号分子识别路径,而在斑马鱼中制作出所谓的“奥德修斯变种”,探明了一对配体和受体组合:SDF-1和Cxcr4b,它们是性腺细胞从发生位置迁移到日后性腺的路标和“盲公棒”。正常的生殖细胞会在斑马鱼胚胎发育时期与其他细胞集中在身体中线,然后从那里开始向将来的生殖腺迁移。而研究人员则为斑马鱼胚胎引入突变,使之成为奥德修斯变种。他们发现,性腺细胞因为细胞膜上的化学趋向受体Cxcr4b发生了变异,不能识别拥有“路标”作用的信号分子,而迷失了方向。它们游离于身体各处。而Cxcr4b也是白细胞游向细菌过程中的重要分子。而在人体和老鼠体内发现,Cxcr4b分子在HIV感染T细胞,B细胞和神经细胞的移动中都起着重要作用。
而免疫学研究则表明,“路标”是SDF-1(stromal cell derived factor 1,中译:基质细胞衍生因子-1),该因子对多种细胞均具有趋向作用[13]。也就是说,SDF-1是Cxcr4b的配体。研究人员为了证实这一点,除了观察正常胚胎之外,还制作了两种变种:第一变种在性腺细胞的必经之路上SDF-1浓度被降低,性腺细胞又成了“奥德修斯”。而第二种变种则是将SDF-1分子散播在斑马鱼身体的其他部位,观察性腺细胞迁移出错。


而研究人员还发现SDF-1对斑马鱼体侧线的细胞迁移有关。鱼类通过一些细小的感受器—神经丘(neuromast)[14] 感受水流变化,这些神经丘排列成线列于鱼体侧皮下,从100多个表皮细胞(被称为体侧线原基细胞,英文:Primodium)分化而成,从耳后基板(Placode)排列直至尾部。它们与体侧线神经相连,通过这些神经传导冲动到中枢再作信号整合。而神经纤维(即神经细胞的轴突)是由神经胶质细胞包绕。因此体侧线原基细胞,体侧线神经纤维和神经胶质细胞是很适合作为研究多种类型细胞共同迁移的模型的。研究人员应用了一种名叫彩色快进分析法(Multicolor-Zeitraffer-Analyse)对斑马鱼的体侧线进行研究。研究发现,三者的迁移是同时进行的。神经胶质细胞跟着神经纤维前进,但神经纤维即使没有胶质细胞的陪伴,也能找出正确的路径。而神经纤维则是跟着体侧线原基细胞前进。
体侧线原基细胞也是透过Cxcr4b分子的帮助去识别路径分子SDF-1。原基细胞的Cxcr4b如果发生变异,细胞不能前移。神经纤维即使正常,也只能因为“领路人”而停步。但是神经纤维即使有变异的Cxcr4b,却也能跟着原基细胞前进。总结说来,原基细胞“拉着”神经纤维行进。马克思·普朗克研究所这项工作也是对很早就已被提出的“神经纤维(轴突)跟着它所支配的感受器迁移”假设的首次证实。这种“拖拉”机制不但在胚胎发育阶段,也在动物的长成阶段,保证了神经纤维与感受器的正确连接。
至于神经胶质细胞,研究人员通过制作一系列的变异,筛选出其中那些神经胶质细胞不能跟随神经轴突一起移动的变种并加以研究,并且对ErbB受体施以阻断剂,发现神经胶质细胞的迁移涉及到神经调节蛋白—Erb信号传导通路。神经胶质细胞膜上具有神经调节蛋白受体ErbB2和ErbB3,它们都是属于受体酪氨酸激酶家族。该家族的成员接受表皮生长因子(Epithel growth factor)或其他肽类的信号,然后两两结合,相互激活再发挥作用。ErbB3能结合配体,与另外的ErbB3或其他家族成员结成同二聚体(Homodimer)或异二聚体(Heterodimer)。ErbB2不会与配体结合,正常情况下只会与其他家族成员组成二聚体(但若果ErbB2过度表达,则会形成同二聚体,与癌症产生相关)。
受体酪氨酸激酶信号传导过程如下,当单体膜外段与信号分子结合后会两两配对,称二聚化,双方催化中心的附近激活唇(activation lip)一或多个酪氨酸残基被磷酸化激活,接着双方会发生构象改变,结果是对ATP亲和力的增高,或是对某种蛋白质结合能力增强,并磷酸化激活受体胞质内其他酪氨酸位点。这些被磷酸化的酪胺酸通常会与适配器蛋白作用。一如其名中的“适配器”,适配器蛋白本身并不具有催化活性,只是为两种酶提供一个沟通的接口,正如日常转插座所起的作用一样。通过适配器蛋白Ras激活蛋白(其实就是Ras的鸟嘌呤交换因子)被激活,它会将Ras-ADP的ADP换为ATP,Ras被激活,信号进一步被传递。在斑马鱼的神经胶质细胞,这条被激活的信号通路会让细胞跟随轴突前进。这种轴突—神经调节蛋白—神经胶质细胞机制保证了胶质细胞与神经纤维的紧密结合[7]。而神经调节蛋白代谢在很多癌症的演进过程中起到重要的作用,异常的ErbB受体信号与肿瘤的发生、发展有着密切的关系[15]。因此,ErbB受体阻断剂有可能成为一种癌症治疗手段。
极化
当细胞胞膜上的受体接触到周围环境里面的迁移信号分子之后,细胞内部与细胞迁移有关的物质会重新分布,细胞显出“前”和“后”两端,为迁移做准备。物质如β机动蛋白的mRNA,Arp2/3复合体还有一些化学感受器,会呈现出前多后少的分布状况,与之相反的是Ca2+。虽然两种物质有着不同的分布趋势,但是它们的目的都是一样的,就是促进细胞的移动性。
β机动蛋白的mRNA在细胞收到迁移信号之后会集中在细胞前端,为这里提供微丝单体。而为Arp2/3复合体七条肽链编码的mRNA也会齐集在突出端。这种mRNA的局部汇聚反过来却需要微丝还有微管的协助。这种mRNA汇聚的发生对细胞迁移有极大的好处。一方面降低了细胞对之调控或降解的难度,另一方面它们的新生翻译产物互相靠近又有利于复合体的快速组装(这种现象被称做共同翻译组装,英文:cotranslational assembly),同时也保证了各自的正确折叠,避免与其他细胞成分发生负面的相互作用[16]。
钙离子的分布与上面提到的不同。Ca2+前端分布浓度低,而尾端高。如果在一迁移中的白细胞侧边放入迁移信号物,人们发现细胞内的钙离子水平先是总体升高,然后再分布成前低后高的状态,细胞转弯。如果此时撤销新放入的信号,白细胞会接着按照新的信号物浓度继续其迁移。很多微丝结合蛋白,如I和II型肌球蛋白,凝胶溶素,辅肌动蛋白(辅肌动蛋白)和丝束蛋白(fimbrin)都受到钙离子的调控。因此钙离子是细胞由溶胶变凝胶(sol-to-gel)过程中重要的一员。前端盖离子浓度低,能激活I型肌球蛋白,抑制微丝分解蛋白,拨转由钙离子调控的微丝交联蛋白,因此有利于微丝网络的形成。而尾部高钙离子浓度则会导致微丝的解聚,激活凝溶胶蛋白,促成溶胶状态的出现,而且会激活II型肌球蛋白,使外皮微丝网络收缩。总的说来,钙离子的胞内浓度梯度对微丝的周转起到了很重要的作用。
运动的四个步骤
当人们观察角膜细胞的迁移时,可看到细胞体外形的改变。这可以类比于人的步行过程,首先是确定前进方向,然后是重复一系列动作循环,即一只脚先往前踏出,并且在地上踏实,而鞋纹则有防止向后滑的功能,再是上身重心前移,最后是后脚提起并向前脚靠拢,完成一个循环。而其中的每个步骤,都受到一个精巧的调节网络控制,以保证原料和能量的合理利用和细胞迁移的有效进行。这四个步骤的示意图请见插图1。
细胞前端突出

这是上面说到的步行过程中前脚伸出。细胞首先会伸出极状足,极状足有两种,一是板状伪足(lamellipodia),另一则是丝状伪足(filopodia),这两种伪足可同时出现在一个细胞膜的某一局部。但构成两者的微丝蛋白结合蛋白有异,板状伪足因为有Arp2/3的帮助,微丝会形成网状,而在丝状伪足里面,微丝成直线状。所以在外形上板状伪足显得扁平,丝状伪足则是薄薄的针状突出。但两者的形成机制究竟是否有别,则仍未有定论。极状足的内部,是因为个体分子的不断加入而变长的肌动蛋白纤维(英文:Actinfilament,即微丝),这个过程被称为多聚化())。而且这些变长的微丝之间会靠一些蛋白,如细丝蛋白的连接而成网成束,加强其韧性。
分布在细胞前端的肌动蛋白多聚化,可以被看作是一种动力来源,不断推细胞膜向前突出。这种被称为“基于肌动蛋白多聚化机制”(actin polymerization–based mechanism)可以理解为不断多聚变长的微丝在内部不断地“顶”着细胞膜往前,一如精子的顶体反应。其过程如下:肌动蛋白单体被不断添加到微丝近胞膜一端,而Arp2/3则为微丝网络不断添加侧枝,促成新的侧枝以70°生成展开,让更多的单体添加到网络上。蛋白质如细丝蛋白(Filamin)则负责巩固这种结构。前纤维蛋白将从多聚体脱离下来的ADP肌动蛋白转换成ATP肌动蛋白,使之从新具备多聚化的能力。而弹性布朗棘轮模型(elastic Brownian ratchet model)则进一步解释突出是如何被不断变长的微丝“鞭”出来的。根据电子显微镜观察得出的结果,微丝的尾部是抵着细胞膜的,肌动蛋白单体无法被加载。但是热能会促使微丝弯曲,这时单体就有机会加入到多聚物的末尾。微丝和一根塑料棒有着相当的刚性,弯曲所保存的势能会帮助微丝变会原来伸直的状态。数量众多的微丝这样“鞭”打细胞膜,会提供足够的动力促使细胞膜突出。
微丝的核化除了Arp2/3复合体外,还需要形成素(formins)中的mDia1和mDia2的协助。而Arp2/3会受到WAVE/Scar, WASP(欧德里症候群蛋白)和N-WASP蛋白的调控。这些蛋白如果出现突变或者功能缺损,患者很有可能会得到欧德里症候群(Wiskott-Aldrich syndrome,或译威斯科特·阿德里希综合征)。WAVE/Scar是Abi,NCK连接蛋白(NCK-associated protein,简称Nap125),特异Rac1连接蛋白(Specifically Rac 1-associated proteins, 简称Sra-1)和HSPC-300多聚体的一部分。这个复合体受小GTP蛋白Rac的调控,Rac会促进Abi, Nap125和Sra-1与WAVE分离,使之激活。WASP和N-WASP则是受到Cdc42调控。这些蛋白质又会受磷酸化过程或磷酸肌醇,WIP和TOCA等分子的控制。
与Arp2/3促成侧枝不一样,形成素会结合到微丝的末端,使之线形延长。形成素也受到小GTP蛋白调控,mDia1受RhoA,而mDia2则受Cdc42调节。它们还需要与前纤维蛋白相互作用才能达到促进微丝多聚化的目的。前纤维蛋白的行为模式可能与抑制微丝末端“帽子”的形成有关,这种蛋白也为Ena/VASP介导的微丝末端多聚化所需。前纤维蛋白和胸腺嘧素4都是G肌动蛋白结合蛋白,而胸腺嘧素4是前纤维蛋白的拮抗剂。但是前纤维蛋白能够与不同的肌动蛋白结合,胸腺嘧素4则不能。所以后者被视为是维持胞质G肌动蛋白水平的重要一员,使之形成G肌动蛋白库,G肌动蛋白可以从中靠前纤维蛋白的帮助进行多聚化。[17]
突出与底质的粘着
该步对应于人的前脚固定。不论细胞的前端是如何向前伸出的,但是伸出后的极状足需要在底质上固定。在显微镜下可观察到肌动蛋白束在细胞前端内部往落点上固定,并发展为一块具有一定结构的斑,被称为“粘着斑”。粘着提供摩擦力,能使细胞巩固向前迈出的步伐而不致向后“滑”。粘着斑的聚合和解聚受到细胞外物质的影响,同时也影响着细胞的行进。
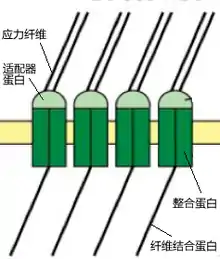
粘着斑由整合蛋白(Integrin)和负在上面的适配器蛋白(Adaptor protein),再加上纤维结合蛋白(Fibronectin)共同构成。整合蛋白是细胞膜上主要的细胞外基质受体,在一系列细胞生理过程中起作用[18]。根据一些实验的结果,带有缺陷的整合蛋白无法使细胞留驻在底质上,但不影响细胞突出的形成。适配器蛋白可看作是应力纤维连接整合蛋白的接口。而整合蛋白则是跨膜蛋白,最终负责直接与外界底质接触的是纤维结合蛋白。它们的结构示意图请看插图11。
不难想象,细胞前端不断生成粘着斑,后端不断解聚粘着斑以供前端使用。但其实前端的粘着斑一样会不断解聚,解聚所得的蛋白质最终由蛋白酶分解,分解产物会被运输到前端的其他位点被再投入使用,称作“周转”(Turnover)。所以前端粘着斑有着两种命运,或是解聚,或是被巩固增大。在活体里面的细胞的粘着斑相比起那些体外培养的,在二维平面上移行的细胞的要小。新生的和早建的粘着斑在结构上差异不大,但成分有不同之处。踝蛋白(Talin)连接整合素和肌动蛋白。有很多报道称,除了肌动蛋白,还有其他信号分子会连接到整合素上,调节整合素和细胞骨架的活性。例如,斑联蛋白(zyxin)会连接到α-辅肌动蛋白(α-actinin)并调节邻近微丝末端上的Ena/VASP活性。虽然粘着斑的初始化(也被称为“核化”)和调控机制是目前细胞生物学研究的热点之一,但人们还是未知道,究竟什么机制决定一块粘着斑是被巩固还是被解聚。可以肯定的是,结合在细胞骨架上蛋白质的共价修饰(covalent modification)形式之一——酪氨酸残基磷酸化是粘着斑形成过程中的重要现象。另外,Rho-GTP酶也被肯定是其中不可忽视的一员。但它本身又会被与粘着斑相关的信号分子所调节,如复合体粘着斑激酶(Focal adhesion kinase,简称FAK),Src,桩蛋白(paxillin),Crk,CAS,PAK和GIT。其中,FAK,桩蛋白还有张力蛋白(tensin)这三种蛋白,都是目前该调节通路“主角”的候选人。其作用方式,现以粘着斑激酶为例以作说明:在细胞粘着斑形成处,整合蛋白会吸引一批蛋白质到自己周围,形成一复合体,以便启动其对其它蛋白质的调控过程。粘着斑激酶是粘着斑形成早期“被招募者”之一,它控制着一大批下游反应,会合诸如适配器蛋白生长因子受体结合蛋白2(Grb2,全称:growth factor receptor-bound protein 2)和磷脂酰肌醇-3激酶(PI3K)的p85亚基起作用。而研究表明,整合蛋白可以通过这条FAK信号通路作用,增加细胞的移动能力,求生(Survival)能力,甚至是癌细胞的转移能力。FAK也因此成为了癌症研究的一个对象[19][20]。過去被認為與發炎反應及細胞凋亡有關的c-Jun氨基末端激酶(JNK;c-Jun amino-terminal kinase),也參與了細胞遷移,它能透過將樁蛋白(paxillin)磷酸化,進而影響粘著斑形成[21]。
值得一提的还有微管在周转过程中发挥的作用。20世纪70年代,瓦斯利夫(Vasiliev, J.M)和他的工作小组用噻氨酯哒唑处理了金鱼的成纤维细胞,细胞内的微管被破坏。这时细胞会失去极性。后来他们提出了微管在细胞迁移中的作用:“稳定住细胞边缘活动的和不活动的部分。”后来,科学家再发现,微管会伸展到细胞前进端早期形成的粘着斑处,所以他们认为微管能稳定细胞与底质的粘着。但是后来的发现是,微管对粘着起到的作用是负面的。微管会限制粘着斑的形成,并且会促进后者在细胞的其它部位被重新利用,后面这种现象被称为粘着斑的周转。等到研究技术允许作活细胞观察的时候,人们才终于认识到,微管与粘着斑的相互作用并非如先前那样认为的少,而是非常的频繁。科学家后来利用隐失波显微术(evanescent wave microscopy)观察得到的结果同样证实了这种观点的正确性。
细胞体前移

当细胞前端固定好之后,细胞的主体就可以前进了。但是这个主体推进的过程细节并不明了。现认为,细胞核和细胞器是被“系”在细胞内部纵横交错的细胞骨架上的,再通过II型肌球蛋白的不断运动,使得这些“货物”得以不断被往前拉。这种观点得到一些实验的证实。例如,科学家发现II型肌球蛋白会分布在细胞伪足和细胞主体的分界线上,这样的分布有利于肌球蛋白的“拉”行为。这个过程受到小Rho GTP酶Cdc42, Rac和RhoA的调控。而这些蛋白相互之间却是拮抗的关系。RhoA 激活Rho-激酶(又名ROCK),后者在激活状态下会将肌球蛋白轻链磷酸酶(myosin light chain phosphatase,简称MLC磷酸酶。该酶为MLC去掉磷酸基团)磷酸化,就是使它失活了,导致细胞的收缩增强。Cdc42也是通过MRCK起到类似的作用。与它们相反的是,Rac会激活PAK,PAK能磷酸化MLC激酶,使之失活,后果是细胞收缩力减退,扩展受阻。但是PAK也能直接磷酸化MLC,增加细胞收缩能力。究竟是哪种作用占优,取决于PAK的空间分布和它的活性调节水平。PAK还会通过激活PIX/PAK复合体调节细胞极化,该复合体在由G蛋白偶联受体所引发的细胞迁移中会分布到细胞前端并发挥其作用。PAK还能通过癌蛋白18(stathmin)的磷酸化作用于微管,减少后者变动(catastrophe,即微管从延长状态转变为缩短状态这个过程)的机会。封闭微丝末端的封闭蛋白,如凝溶胶蛋白则主要靠磷酸肌醇调节。而切断微丝的蛋白:丝切蛋白则受到LIMK介导的磷酸化影响。而LIMK又会受到PAK介导的或是Rho-激酶介导的丝氨酸/苏氨酸磷酸化的调节。[17]
牵引尾部往前
最后一步,类似于人步行时后脚往前收。细胞后端,或称为尾部与底质分离并被牵往前方。这个过程可能与应力纤维的收缩,或是简单的细胞弹性变形有关。值得注意的是,细胞后端是被“撕”(ripping)离底质的。就是说,细胞的尾部会在底质上留下一部分细胞膜残片。而尾部的粘着斑也会卸载,否则细胞收缩会撕开细胞。这个卸载的过程包括,粘着斑部件以胞吞作用的形式被内吞,然后会被发动蛋白承载并回收转运,这个过程有微管的参与。在肌球蛋白介导的胞体收缩过程中,RhoA和Rac通过Rho-激酶和PAK可以调节MLC的磷酸化,这也为尾部粘着斑解聚做出贡献。还有,尾部磷酸酶,如钙调磷酸酶(calcineurin)的磷酸化作用或蛋白质被蛋白酶降解,如踝蛋白被钙蛋白酶(calpain)降解都会下调粘着斑与底质的亲和力。钙离子可能在此过程中会调节神经钙蛋白和钙蛋白酶[17]。
与细胞迁移有关的生理过程
细胞迁移是多种生理过程的前提,例如创伤恢复,神经嵴细胞的移行,急性炎症中白细胞的渗出还有癌细胞的转移。
胚胎發生

高等动植物成体的结构非常复杂,但都是来自于一个受精卵。受精卵不断分裂,所得出的细胞会移动,还会通过基因的开启或关闭进入分化途径,形成特异的细胞,执行其被指定的功能。胚胎發生(Embryogenesis)通常被分为三个阶段。第一阶段是原肠胚形成,指受精卵分裂到囊胚后,经过囊胚的折叠逐渐成为有三层胚层结构的原肠胚的整个过程,此过程后身体各部分的构造方向已基本定下。第二阶段是器官形成(Organogenesis)。最后阶段则是各器官继续成熟完善至成体状态。
动物的胚胎发生涉及大量的细胞迁移行为。科学家在研究这些细胞迁移时,可以使用无毒性的染料,或者是可以遗传的遗传学标签(heritable genetic label)对那些将要迁移的细胞进行标示,以追踪其动向。例如,科学家可以从鹌鹑胚胎内取出其将来要成为翅膀的体节(Somite),将该体节移植入培养了两天的小鸡胚胎中。经过一周后,将小鸡翅膀部分剖开并观察其肌肉,可见它们是来自鹌鹑体节的。
观察无脊椎动物如海胆(sea urchin)的原肠胚形成过程(Gastrulation),即从囊胚(Blastula)到原肠胚(Gastrula)的形成过程。开始时囊胚是由约1000个细胞组成的,球状中空结构,球腔即为囊胚腔(blastocoele),球壁由单层细胞组成,植物极(Vegetal pole,可以看作是日后海胆身体的尾部)上皮比较厚,出现细胞内馅的倾向。然后一些间充质细胞开始游离植物极上皮,并在囊胚腔内爬行。而植物极上皮也开始向内折叠,上皮外围的细胞会向囊胚腔伸出丝状伪足,帮助上皮向动物极(可以看作为海胆日后的头部)移动并最终与囊胚腔另一端接合。上皮内陷折叠时,留下的空隙会成为日后的肠道,上皮与囊胚壁接触之处则是日后海胆的口部。
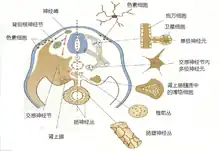
脊椎动物的神经嵴细胞,在胚胎期会不断从背侧向腹侧移行。其中一部分移行于外胚层下方,将来会分化为色素细胞,而那些行走的稍深一点的细胞,会形成后来交感神经的神经节细胞,肾上腺髓质。而颈部和骶部的神经嵴细胞则会沿着身体纵轴移到肠壁。就是说,日后的组织肠神经丛,神经节神经元,肾上腺的嗜铬细胞(chromaffin)都是由神经嵴细胞迁移分化得出的。
值得注意的是,在此过程中,沿途的不迁移细胞可能会影响迁移细胞的行为,改变它们的去向,甚至决定迁移细胞是否能存活。同来自神经嵴的性细胞,血细胞前体和色素细胞都受到一种Kit—Steel因子机制的调节。Kit是一种跨膜受体,其配体是Steel因子。沿途的细胞或者是终点处的细胞会表达Steel因子,激活迁移经过的细胞上的Kit受体。而Kit受体的激活是这些细胞存活和增殖的前提。在一个个体中,两者之中的任一者出现突变,患者的体色,血细胞供应和性细胞的形成都会出现异常,例如患者额头可见一白斑。
损伤修复
损伤会对机体造成身体部分细胞和组织的丧失,机体需要对损伤进行修复(repair)。修复有两种不同的形式,再生和纤维性修复。这两个过程都涉及细胞迁移。例如上皮组织中的一种——鳞状上皮如果出现缺损,其边缘和底部的细胞就会分裂增生,并向缺损中心迁移。虽然很多种类的细胞都具有分裂再生的能力,如上皮细胞,但是它们最终能否成功修复受损组织却要依赖于细胞外基质。例如细胞外基质的一种成分—透明质酸(hyaluronan),存在于迁移细胞的周围,它能够抑制细胞间的粘附,促进细胞迁移。
如果损伤伴有炎症的发生,实质细胞即使具有再生能力,也很难独自完成修复工作。这时就要靠肉芽组织(granulation tissue)进行修复了。肉芽组织会首先增生,并溶解吸收坏死组织和异物,填补空缺,再最终转化为瘢痕组织完成修复。肉芽组织由新生的毛细血管和成纤维细胞组成。血管内皮细胞会先迁移到受损部位新生形成血管。接着血小板,炎细胞以及活化了的血管内皮细胞会释放生长因子,如TGF-β,DGF,表皮生长因子,FGF和促纤维化性细胞因子如IL-1和TNF-α等分子,这些因子能吸引单核巨噬细胞和成纤维细胞的增殖和迁移。前者会在受损部位清理细胞外碎片,纤维蛋白和其他外源物质。而成纤维细胞则会合成细胞外基质并不断积聚。最后经过肉芽组织的结构调整,最终形成瘢痕。
细菌感染
致病源微生物对宿主细胞的入侵同样会造成微丝的动态改变。很多致病细菌经过演化,甚至是和宿主共同演化(coevolution),发展出一套生存策略,利用宿主细胞动力蛋白的多聚化,为自身的入侵,繁殖或扩布创造条件。而通过研究这些细菌对细胞骨架的作用途径,科学家可以对细胞迁移的调控作更深入的了解。
1994年研究人员发现,分布在细胞外或是存在吞噬泡(Phagosome)内的格兰氏阴性细菌,如志贺氏菌(Shigella)和沙门菌(Salmonella),演化出一套III型分泌系统(Type III Secretion System,简称TTSS),可以将细菌蛋白质注射入真核细胞细胞质内,模拟细胞内的细胞因子,控制肠上皮细胞或肠内皮细胞细胞骨架的重整理(Rearrangement),赋予这些不具备内吞能力的细胞以胞吞能力,以便自己进入细胞内,这种机制被称为触发器机制(Trigger)。具体地说,沙门菌会分泌蛋白质SipC。SipC的N端会与肌动蛋白结合,C端具有促进肌动蛋白核化的功能。而志贺氏菌有着与SipC同源的蛋白质IpaC,则会激活Cdc42和Rac。这样,肌动蛋白会在细菌与细胞结合处多聚化,为细菌的进入创造条件。同时,志贺氏菌还会分泌IpaA,这种蛋白会结合细胞内的粘着斑蛋白(Vinculin,或译钮带蛋白),并将后者带到细菌与细胞接触之处,形成粘着斑样结构,被称为入侵焦点(Entry Focus)。这种IpaA-粘着斑蛋白能使所在之处的微丝解聚,使得志贺氏菌更容易进入细胞。
另外还有拉链机制(Zipper)。细胞膜表面具有一系列受体,它们是为胞胞连合和胞底质联合所需的。一些细菌,如单核细胞增生李斯特氏菌(Listeria monocytogenes)和鼠疫杆菌(Yersinia)会在自己表面表达出这些受体的配体,这样可以诱使细胞伸出伪足包绕自身,并最终被吞入胞内。李斯特氏菌会表达一种名叫内化素A(Internalin A)的蛋白,它会结合胞胞连合蛋白质E钙粘蛋白(E cadherin)。而耶尔森氏菌则会使用侵染素(Invasin)结合β1-整合蛋白(β1-Integrin)。目前人们认为,这两种途径会最终让肌动蛋白-肌球蛋白组合产生拉力,将细菌拉入胞内。
白细胞渗出
炎症反应最重要的功能是将白细胞送到炎症灶,所以白细胞的渗出是炎症反应最重要的特征。其过程如下:炎症部位的血管内皮细胞会在组胺,白三烯等物质作用下,加上骨架重构,穿胞作用的增强和损伤而收缩,随之而来的是血管通透性的增加,这进一步造成血流速度的减慢甚至是停滞。到达该处的白细胞因此会离开血管的中心部,边集于血管壁,不断滚动直至最后在胞膜表面选择素(Selectin)的作用下与内皮细胞黏附。然后,白细胞会在内皮细胞连接处伸出伪足,以阿米巴运动的方式穿过间隙到达炎症灶,需时2到12分钟。而游出的细胞也有分先后,早期先是中性粒细胞游出,48小时之后再轮到单核细胞。游出的白细胞然后会在炎症灶附近搜索细菌产物,补体成分,细胞因子和白三烯。这些物质能吸引甚至激活白细胞,将白细胞带到炎症部位并发挥其吞噬,免疫和组织损伤作用。中性粒细胞和巨噬细胞能吞噬病原体或组织碎片,而巨噬细胞还会执行其抗原呈递功能,激活B,T淋巴细胞,以杀伤病原体[22]。
癌症转移
目前人们对恶性肿瘤的研究是多方面的,从癌症的产生到转移,血管供给以及分裂增殖都一直是医学和生物学研究的热点。癌症细胞增殖失控,短时间内可以繁殖出大量后代,这样首先会造成生长空间的局促和养分,如氧气的紧张。这样恶性肿瘤内会形成一片坏死区,正如上面在组织损伤里面提到的,机体会尝试“修复”这些损伤。坏死组织会释放出一系列促血管生成因子,如血管内皮生长因子(vascular endothelial growth factor),还会招来巨噬细胞。巨噬细胞也会释放大量促血管生成细胞因子和生长因子。过程中有一类名叫高机动性组蛋白(high-mobility group proteins,简称HMGB)可能起到协调作用。最近研究表明其中的一种:HMGB1能强烈诱导血管内皮迁移[23]。 新生血管既解决了癌组织的供给问题,也为癌细胞的远端转移提供了管道。癌细胞借血道转移到远端器官,并在那里增殖。转移是恶性肿瘤的确凿证据,同时也是癌症患者的主要死亡原因。

癌细胞既能直接蔓延,如晚期子宫颈癌可直接蔓延至直肠和膀胱;它们又可以对周遭组织进行浸润,结果是形成边界不分明的癌组织,或是进入血管转移到远端(见远端转移)。两者的机制比较复杂,目前还有未明的问题尚待解决。目前根据观察可将癌症浸润分为四步。首先癌细胞表面的粘附分子会减少,与周围的细胞彼此分离,被“解除束缚”。同时癌细胞与基底膜的粘附却会增加,这是癌细胞通过增加自身基底面层粘连蛋白(laminin)受体实现的。然后癌细胞会释放蛋白酶,用以降解细胞外基质成分,如IV型胶原酶,使基底膜受损,产生缝隙。最后是癌细胞阿米巴运动样的迁移,钻过基底膜的缝隙,到达底下的间质组织。癌细胞此后会继续用蛋白酶为自己在间质组织开路,直到血管,然后它会以同样方式进入血管,经血到转移后,又以同样方式出管。
癌症转移过程中涉及的信号传递途径,竟然有很多是胚胎發育过程中所必需的。科学家在研究中发现越来越多重要的分子,如N-钙粘蛋白(N-Cadherin),核因子κB(NF-κB),骨连接素(Osteonectin),血栓素A2(Thromboxane A2)和Ras,都是促成癌细胞迁移的因子。
以ras基因(鼠肌小节,Rat sarcoma)这个原癌基因为例,其翻译产物是一21达尔顿重的G蛋白Ras。Ras可分为四种经典Ras蛋白:H (Harvey)-Ras,N(Neuroblastoma,成神经细胞瘤)-Ras,和一对选择性剪接产物K(Kirsten)-Ras4A和4B。从插图2可见,Ras在信号传导通路中位于中心位置,它的变异所造成的严重后果可想而知。人类30%的癌症中被查证有变异的ras基因,它们的产物一方面能抑制细胞凋亡,还会加快癌细胞转移。H-Ras变异蛋白可见于膀胱癌和肾癌。而在几乎所有的乳腺癌中都可看到变异的K-Ras。另外在肺癌,大肠癌和直肠癌中都可见其身影。K-Ras的作用很可能是通过Ras-Raf-MEK-ERK途径实现的。这条途径不但能促进血管生成,还会诱发癌细胞的浸润和转移。
溶鞘磷脂,(Lysosphingomyelin,或称鞘氨醇磷酰胆碱,英文Sphingosylphosphorylcholin),简称SPC,目前被怀疑是癌症转移的启动子。溶鞘磷脂是一种具有生物活性的胞内外信号传导物质,能诱导细胞移行。它先是在患有A型尼曼匹克症(Niemann Pick type A)病人脑部被发现。它的受体是胞膜上的卵巢癌G蛋白偶联受体1(Ovarian cancer G-Protein coupled receptor 1)。研究发现,患有卵巢癌的病人,其腹水中SPC水平明显高于患良性卵巢肿瘤的患者。
在显微镜下通过观察胰腺癌细胞在存在和不存在SPC情况下的活动和胞内角蛋白(Keratin)的分布,可以大概了解SPC在癌症转移中的作用。角蛋白属于中间纤维,它的作用是维护细胞的稳定,赋予胞体一定的硬性。在未经SPC处理的胰腺癌细胞中,角蛋白均匀分布在细胞质中。若它受到SPC的处理,可以观察到其角蛋白会被集中分布到细胞核周围,而细胞膜下角蛋白的浓度则会下降。角蛋白这样的重新分布无疑使得癌细胞外围变得更有柔韧性,让它们更容易通过它们在基底膜打开的狭窄孔洞。
而在1996年被发现的RhoE则饰演着抑制癌症的角色。与其他游移于激活/失活状态的G蛋白不同,RhoE持续与GTP结合,一直处于激活状态,所以其功能的上下调节靠的是其表达水平。在2007年一份研究报告中指出,RhoE在肺癌患者中表达异常,而且还与患者吸烟史有关[24]。根据另一项研究,RhoE不仅在细胞增殖方面起到调节作用,则在细胞迁移方面发挥着其功能,甚至将某些癌细胞引向细胞凋亡[25]。路德维希癌症研究所(Ludwig Institute for Cancer Research)的研究人员发现,RhoE会在生长因子的刺激和DNA损坏的情况下表达增高,因此有可能成为癌症的生物标记。RhoE能阻止肌动蛋白的聚合和应力纤维的形成,因此细胞迁移的能力会减弱。RhoE成为癌症治疗的又一新目标[26]。
参考文献
引用
- Glenn Elert. . hypertextbook.com. [2007-05-14]. (原始内容存档于2007-04-03).
- Douglas A. Lauffenburger and Alan F. Horwitz. . Cell. 1996, 84 (3): 359–369 [2013-11-20]. (原始内容存档于2015-06-13).
- . biophysics.com. [2007-06-04]. (原始内容存档于2007-09-29).
- . dakewe.com. [2007-04-06]. (原始内容存档于2006-11-02).
- . 东胜. [2012-07-10]. (原始内容存档于2012-07-10).
- (PDF). cqvip.com. [2007-09-30]. (原始内容 (PDF)存档于2007-09-30).
- Gilmour, D. Knaut. (PDF). 2006.
- 董理;彭谨;杨明亮. . wanfangdata.com.cn. [2007-05-02]. (原始内容存档于2007-09-30).
- P Maurel, U Rauch, M Flad, R K Margolis, and R U Margolis. . pubmedcentral.nih.gov. [2013-11-20]. (原始内容存档于2019-03-13).
- Gopalakrishna, R. . pnas.org. [2013-11-20]. (原始内容存档于2013-11-20).
- . ncbi.nlm.nih.gov. [2013-11-20]. (原始内容存档于2013-11-20).
- . .ncbi.nlm.nih.gov. [2013-11-20]. (原始内容存档于2013-11-20).
- (PDF). cqvip.com. [2007-09-30]. (原始内容 (PDF)存档于2007-09-30).
- . zipcodezoo.com. [2013-11-20]. (原始内容存档于2012-03-10).
- 郭晓宁 林莉萍. . cqvip.com. [2013-11-20]. (原始内容存档于2007-09-28).
- Moore, M. J. . SCIENCE. 2005, 309 (SPECIAL ISSUE): 1518.
- Vicente-Manzanares M, Webb DJ, Horwitz AR. . J Cell Sci. 2005, 118: 4917–4919 [2007-03-31]. PMID 15888700. (原始内容存档于2007-02-18).
- Guan JL. . Int J Biochem Cell Biol. 1997, 29: 1085–96. PMID 9416004.
- M. Ginsberg; J. Ladbury. . biop.ox.ac.uk. [2013-11-20]. (原始内容存档于2011-07-12).
- Zheng. . stjude.org. [2007-05-14]. (原始内容存档于2005-03-13).
- Cai Huang; et al. . Nature. 2003, 424: 219–223. PMID 12853963.
- Rashmi Yadav, Karen Y. Larbi, Rebecca E.Young, Sussan Nourshargh. . schattauer.de. [2013-11-20]. (原始内容存档于2013-11-20).
- SCHLUETER, J.; WEBER, H (2006). . cat.inist.fr. [2013-11-20]. (原始内容存档于2013-11-20).
- Cuiyan Z, Jie H. . Cancer Biol Ther. 2007. PMID 17312384.
- Poch E, Minambres R. . Exp Cell Res. 2006, 313 (4): 719–31. PMID 17182035.
- Ludwig Institute for Cancer Research. . licr.org. [2009-06-27]. (原始内容存档于2009-06-27).
书籍
- Wedlich, D(2005): Cell Migration in development and disease, Wiley-VCH Verlag GmbH & Co.KGaA, Weinheim ISBN 3-527-30587-4
- Lodish, H.; Berk, A. (2001): Molekulare Zellbiologie, 4. Auflage, Spektrum Akademischer Verlag GmbH Heidelberg, Berlin ISBN 3-8274-1077-0
- Alberts, B. (2004): Molecular Biology of the Cell, Garland publishing, Taylor & Francis Group
- 田润刚(2004):《细胞生物学教程》,第4版,绍兴文理学院生物科学系
- 李玉林主编(2003):《病理学》,第六版,人民卫生出版社
- Gilmour, D.; Knaut, H.(2006): Zellwanderungen im Embryo des Zebrafisches: Wie Zellen ihren Weg beim Aufbau von Organen finden. Max-Planck-Institut für Entwicklungsbiologie, Tübingen
- Johmann, K.(1998): The Computer inside you, 4th Edition